トンボの生態学
5.幼虫の生活 目次
トンボの生態学 5.幼虫の生活
幼虫の生活する水環境
トンボの幼虫(通称ヤゴ)は,ほぼ例外なく水中生活をしています.特に日本産のトンボ幼虫はすべてそうです.サラサヤンマやムカシヤンマなど,一部のトンボ幼虫が水のないところで見つかったりしますが,これらも飼育環境下ではずっと水中で生活しています.ムカシトンボは,羽化に先立って水中から出て陸上で過ごす時期があります.しかしこれも成長のほぼすべての時期を水中で生活しています.したがって,水環境はトンボ幼虫の生活のさまざまな面に影響を及ぼすことになります.
Ⅰ.呼吸器官と溶存酸素量
(1-1) 呼吸器官
トンボ幼虫が水中で生存していくにあたって,その生死に関わるもっとも大きな水質条件は「溶存酸素量 DO*1」です.ここでは溶存酸素量について考える前に,まず呼吸器官について理解をしておきましょう.
昆虫の呼吸器官については中学校の教科書にも掲載されていましたが,呼吸は気管系で行われています.これは,気門から取り入れた空気 (酸素) をそのまま気管によって全身に運ぶシステムで,人間のように血液によって運ぶシステムとは違っています.昆虫はものすごい周波数で羽ばたくので,その筋肉の動きを支えるための酸素供給は体液による運搬では間に合わず,直接空気を送り込むシステムを完成させたと考えられます.
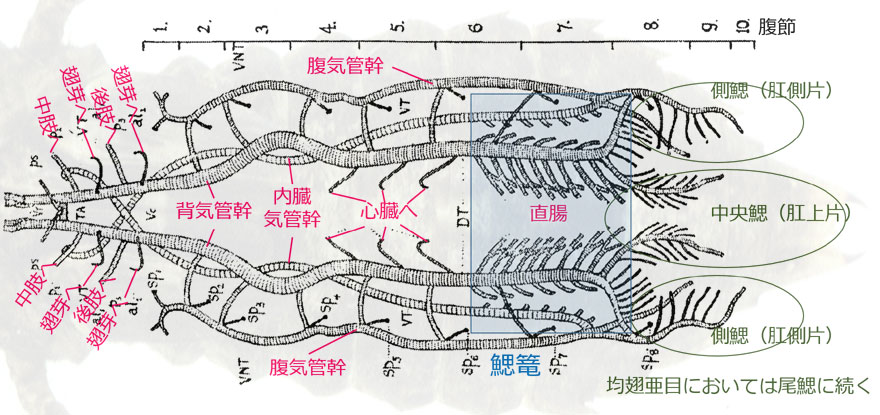
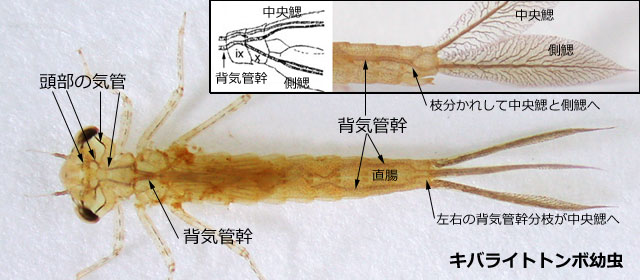
トンボの成虫は陸上生活をしているので気門から空気を取り込むのはたやすいですが,水中生活をしている幼虫では少々大変なことになりそうです.不均翅亜目とムカシトンボ亜目の幼虫は鰓呼吸をしています.鰓は直腸にあり直腸鰓またはその形態から鰓篭 (さいろう) と呼ばれています.肛門から水を取り込んで,この鰓で水に溶けた酸素を取り込み二酸化炭素を水中に排出して,ガス交換をするわけです.気管系の解剖学的な研究は100年以上も前に行われていて,それらは Tillyard (1917) にまとめられています(図1).均翅亜目では太い気管が尾鰓につながっているようすが直接観察できます.尾鰓には細かい毛細気管が全面に分布しており,重要なガス交換の器官になっているのが分かります(図2).
図1.不均翅亜目 Dendroaeschna conspersa 幼虫の気管系(頭部を除く).直腸において,内臓気管幹,背気管幹から多数が枝分かれしている部分が直腸鰓といわれるもので,全体をさして鰓篭と呼ぶ.背気管幹の後先端部は外側に屈曲し,腹気管幹と内臓気管幹の合流した気管とさらに合流する.なお「腹気管幹」の和名称は見つからなかったので筆者が付けた.Tillyard (1917) に追加.
図2.キバライトトンボ幼虫の生体に見られる気管系.普通に観察しても均翅亜目の太い気管系は体表を通して透けて見える.基本的には図1の不均翅亜目と同じである.背気管幹が後方で分枝し,中央鰓と側鰓に接続しているのが分かる.そしてこれらにはたくさんの毛細気管が分布している.この写真から見て,尾鰓がガス交換の器官であることが明らかだろう.手書きの図は Tillyard (1917) を改変.
体を前後方向に貫くもっとも大きな気管が一対の背気管幹で,これは頭部にまで延びていて,途中で,心臓,脚,翅芽へと分岐しています.背気管幹の尾部は直腸部で細かく枝分かれし,鰓篭を形成しています (図1).さらに尾部の末端は,一度分枝をしたあと,腹気管幹と内臓気管幹が合流した気管と合流し,その末端は細かく枝分かれしています.これらの部分は,均翅亜目においては尾鰓へと伸びます (図1,図2).
背気管幹以外には,内臓気管幹,腹気管幹が前後方向に伸びています.腹気管幹には成虫の腹部第1~8節に存在する気門(sp
1~sp
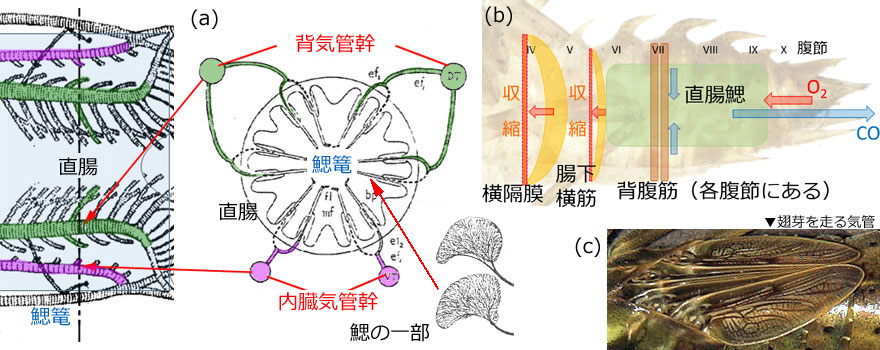
8)への分枝がありますが,幼虫ではこれらの気門は閉じています.また6本の気管で背気管幹とつながっています.内臓気管幹の後部は細かく枝分かれし背気管幹の分枝とともに鰓篭を形成しています (図3(a)).
図3.(a) 鰓篭部分の断面図.背側に位置する左右の背気管幹から枝分かれした一対の気管 (緑色),および腹側に位置する内臓気管幹から枝分かれした気管 (桃色) が,直腸の壁を越えて鰓を形成している.鰓の一部を拡大すると毛細気管が密に枝分かれしているのが分かる.なお鰓篭の形態は,上図を含め発生学的に3つのパターンがあるという.Tillyard (1917) を改変.(b) 直腸における水の吸排水を実現する筋肉の模式図.(c) 不均翅亜目の翅芽に見られる気管系 (オオルリボシヤンマ).すでに気管に沿って黒い成虫の翅脈が形成され始めている.翅脈形成前は白い気管が見える.翅芽もガス交換に役立っていると考えられている.
ガス交換は,横隔膜と腸下横筋および各腹節にある背腹筋によって,肛門から吸排水を行って実現しています (図3(b)).横隔膜と腸下横筋は後方に反った形の筋肉で,これらが収縮すると反りがまっすぐになって直腸を引き (赤短矢印),それによって肛門から水が入ってきます (赤長矢印).また排水の時は横隔膜と腸下横筋が弛緩して後方に反り,背腹筋が縮んで (青短矢印) 直腸を押し込み,肛門から水を押し出します (青長矢印).一般に吸水に比べて排水の方が勢いがあります.これは二酸化炭素を多く含む水をできるだけ遠くへ排出するためです.吸水の時は肛門の開きを大きくし,排水の時は肛門の開きを小さくすることでこの違いを実現していることが,ルリボシヤンマで知られています.またこの吸排水運動の頻度は,低酸素下では増加することがいろいろな種で確認されています (Corbet, 1999).
均翅亜目には直腸にはっきりとした鰓構造がみられません.カワトンボ科の Calopteryx splendens では,直腸壁に毛細気管が発達した小さな突起があることが知られています (Miller, 1993).また上述のような直腸のポンプ運動も見られます.横隔膜や腸下横筋がなく,吸水は別のしくみで小刻みに行われるようです.また排水は一気に行われます.このポンプ運動は,直腸の内壁に分布する毛細気管を通して,ガス交換に役立っていると考えられています.
均翅亜目では,尾鰓,翅芽,体表面でのガス交換が大きな割合を占めているものと思われます (Corbet, 1999).尾鰓には細かく枝分かれした毛細気管が分布しており,ガス交換に有利な形状をしていますが (図2),水温が上がるなどして酸素濃度が低下すると,尾鰓を左右に振るような運動をします (図6).一方尾鰓が取れても幼虫は生きていますので,直腸内壁および翅芽 (図5) や体表面でのガス交換も重要な役割を持っていると考えられます.例えば翅芽がガス交換を行っている状況証拠として,流水性種のミヤマカワトンボをバットに入れておくと,翅芽を開くような姿勢が見られます (図4(a)).他の均翅亜目の幼虫でもときどき見られます(図4(b):オツネントンボ亜終齢幼虫).
図4.(a) 流水性種のミヤマカワトンボ幼虫(おそらく酸素要求性が高い)をバットに入れて太陽光の下に置いておくと,やがて翅芽を開くようになる.これは水温が上がって酸素濃度が低くなり,酸素の取り込みを高めるための姿勢であると思われる.(b) オツネントンボの亜終齢幼虫の翅芽も目一杯開いている.
(1-2) 溶存酸素量を決める要因
水中の酸素は,空気中から水面を介して水に溶け込んだり,水草や植物プランクトンの光合成による酸素放出によって供給されます.一方,水生動物や動物プランクトンの呼吸,およびバクテリアによる水中の有機化合物の分解時に消費され,減少します.通常ヒトの手の入らない自然界では,それぞれの水環境に応じて,これらの増加・減少のバランスが取れています.

図5.ハグロトンボの翅芽.スジのように見えるのは翅芽内を走る気管である.

図6.均翅亜目の幼虫を低酸素下におくと,尾鰓を左右に振るような動きをすることがある.これは尾鰓のまわりの水を,新鮮な水と入れ換える運動と考えられている.
ただこのバランスは,同じ水域でもいくつかの要因によって変動します.例えば水温です.一般に気体は水温が上がるとその溶解度が下がることが知られています
*2.また水面の波立ちが大きくなると,より多くの酸素を溶け込ませる要因になります.また水面付近は空気と接していて酸素が溶け込んでおり,底近くは泥中のバクテリアによって有機化合物の分解を行い酸素を消費していますから,水深によって溶存酸素量が異なることになります.
つまり幼虫の生息場所という点からみると,
水温,
水の動き,
水深の3つが,溶存酸素量に大きな影響を与える要素となります.このうち水温に関しては,グローバルなトンボの分布とも関連してきますので,別項で扱います.水の動きに関しては,私たちがもっとも普通に認識できることとして,流水性種と止水性種がかなりはっきりと分かれていることがあります.これは次項で検討します.
この項では水深について見ておきましょう.特に止水域の水底付近は酸素濃度が下がると予想されます.多くの水底で生活しているトンボ幼虫は,酸素不足に遭遇したとき,移動することで対処しているようです.例えば,発電所の冷却用貯水池に生息している北米のサナエトンボ科
Aphylla williamsoni は,加温された場所 (つまり溶存酸素が少ない場所) ではより水面近くに集まっているということが観察されています (Thorp & Diggins, 1982).
サナエトンボ科の幼虫は,一般に底の砂泥中に潜って生活しています.日本産の止水性サナエトンボでは,コサナエ属,ウチワヤンマ属,タイワンウチワヤンマ属の3属が代表的なものでしょう.このうちコサナエ属では,先端の腹節第10節が伸長しているという特徴があります.これは,腹部先端を上方に曲げて泥から出しガス交換をするための適応形態だと思われます.コサナエ属のタベサナエは腹部第10節が少し短いのですが (図9右),この種は流水にも生息している点で,他のコサナエ属の種とは異なっています.また大湖の深みに棲むメガネサナエも腹部第9節が長くなっています.
図8.大湖の深みに棲むメガネサナエは腹部第9節が縦に長い.コサナエ属のオグマサナエは腹部第10節が長い.これらの幼虫は,右のフタスジサナエ幼虫のように,おそらく腹部先端を上方に曲げて泥から突き出してガス交換を行っている.
これらと異なるのが深みに棲むウチワヤンマです.またタイワンウチワヤンマは冬になると深みへ移動するのか,夏と同じ場所では幼虫がなかなか採れません.これらの幼虫は腹部先端は伸びていません (図9).一方これらは肛門から水を噴き出す「ジェット推進」でよく泳ぎます.これらの幼虫は
Aphylla williamsoni のように,移動することで酸素濃度の減少に適応しているのかも知れません.一度,ウチワヤンマの中齢幼虫が,水際でジェット推進して泳いでいるのを観察したことがあります (1993年10月11日).
図9.水深のあるところに生息するウチワヤンマ,冬季に深みに移動する傾向があるタイワンウチワヤンマ,いずれも腹部先端は長くなく,ジェット推進で移動する.タベサナエは,他のコサナエ属に比べて腹部第10節が短いが,流水にも生息する.
(1-3) 流水性種と止水性種
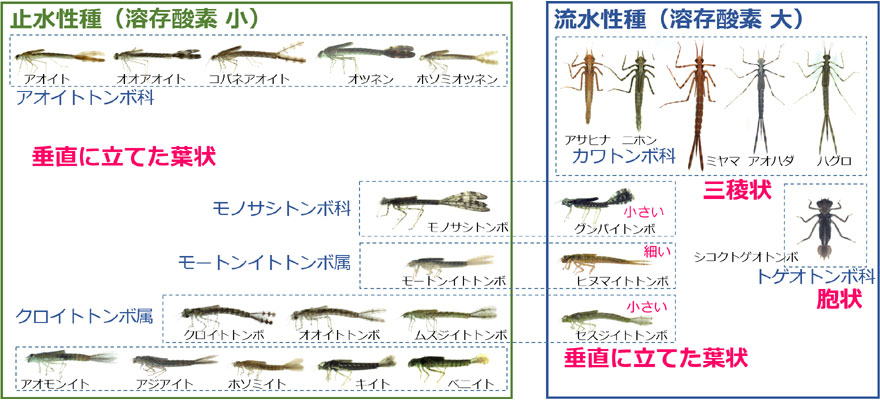
トンボは,河川のような流水に生息する種と,池沼のような止水に生息する種におおむね分かれています.河川は常に水が動いており段差などで曝気もよく行われていて,酸素濃度が高い環境であり,水が動きにくい止水は河川に比べると酸素濃度が低い水環境だと考えられます.図10は兵庫県に分布する均翅亜目にシコクトゲオトンボを加えた幼虫の尾鰓を,流水性種と止水性種で比較したものです.
*2.気体の溶解度は一般に温度が上がると小さくなる.
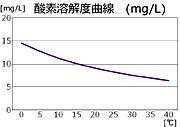
図7.酸素の溶解度曲線.
図10.兵庫県産(トゲオトンボ科を除く)の均翅亜目の止水性種と流水性種の尾鰓形態の違い.止水性の種は垂直に立てた葉状の尾鰓を持っているが,流水性種では,三稜状,胞状で,垂直に立てた葉状のものは,同属・同科の他の種より面積が小さい.
止水性の種はすべて垂直に立てた面積の大きな葉状の尾鰓を持っていて,ガス交換を効率よく行うための形態的な適応だと考えられます.一方流水性の種の尾鰓はさまざまな形態に分化していて,垂直に立てた葉状以外に,三稜状,胞状などのものがあります.また同じ科や属内に止水性種と流水性種が含まれている場合,流水性種の尾鰓の方が面積が小さい傾向があります.
セスジイトトンボはクロイトトンボ属の中で尾鰓がやや小さいだけですが,これは止水にも生息している種です.グンバイトンボは池で羽化殻が見つかったことがあります (東・青木,2025).ヒヌマイトトンボは流水性といっても河口近くの非常に緩い流れの場所に多く生息しています.これらは面積は小さいとはいえやはり垂直に立てた葉状尾鰓を持っていて,止水(的)環境でも生活できるようになっていると考えられます.
少し話は違いますが,アジアイトトンボは,越冬中の幼虫の尾鰓は細長く,春に孵化し夏に羽化する二化目の幼虫は尾鰓の幅が太くなります.これも,冬は低温で溶存酸素が高く,夏は高温で溶存酸素が低いことへの適応かも知れません.
次に流水性種について見てみましょう.シコクトゲオトンボの胞状尾鰓は,この科が水が表面を流れるような岩に貼りついて生活するために,吸着する役割を持っていると考えられています (欄外図11).これら以外にも,ハナダカトンボ科の尾部付属器 (欄外図12) など,これらとは異なる尾鰓を持つ種がいます.
いずれにしても,止水性種が垂直に立てた葉状一択であるのに対し,尾鰓がいろいろな形態・機能に分化しているのは,流水の酸素溶存量が多く,体表,翅芽,直腸の表面だけで十分な酸素を取り込めるからだろうと思われます.逆に言えば,尾鰓がガス交換の器官として特化していないため,流水性種は止水では生活できないのかも知れません.実際,止水性種幼虫が流水環境に見いだされるのはよくあることですが,逆のケースはあまりありません.
これら特殊な形態の尾鰓がどれほどガス交換に役立っているかは分かりませんが,例えば図4で,ミヤマカワトンボの三稜状尾鰓は大きく開いており,アサヒナカワトンボも止水に入れておくと同じように翅芽と尾鰓を開きますので (欄外図13),やはりガス交換を行っていると推測できます.均翅亜目には,これら以外に,ミナミカワトンボ科にある腹側鰓と呼ばれる糸状の呼吸器官など (欄外図14),これらとは異なる構造を持つ種がいますが,これらについては,種特異的となることもあり,ここでは検討しません.
不均翅亜目については,サナエトンボ科の多くの種や一部のヤンマ科の種,オニヤンマ科,ミナミヤンマ科,ヤマトンボ科,一部のエゾトンボ科などに流水性種がいます.ムカシトンボ亜目のムカシトンボも流水性です.しかしこれらには止水性種と比べて目に見える外形的な相違が見られないため,なぜ流水を好むのかについて比較検討するのは,特別な工夫をして調べない限り困難でしょう.
不均翅亜目に関して,実際に観察できることで興味深い現象は,止水性の種が南方の水温が高いところへいくと,流水に入り込むことが多いということです.例えばベニトンボ幼虫は,沖永良部島では流水環境でたくさん見つかりましたが,最近北上してきた近畿地方の個体はほとんど止水環境に見いだされています.また琵琶湖のような大湖に,流水性種が割合多く見られることがあります.大湖では風波が生じますので,特に波打ち際では流水環境と同じように,酸素がより多く溶け込むことが考えられます.これが流水性種が大湖で生活できる要因でしょう.なおこれら流水性種成虫が大湖を自らの生息場所と判定するのは,長く直線的に続く水際線が,川と同じように見えるからだという説があります (Corbet, 1962)
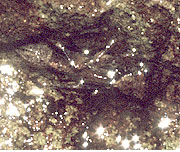
図11.トゲオトンボ科 ヤンバルトゲオトンボは表面を水が流れる岩に貼りついて生活している.

図12.ハナダカトンボ科 ヤエヤマハナダカトンボの尾鰓(尾部付属器).側鰓は堅く棘状で,中央鰓は肛上片様になっている.

図13.アサヒナカワトンボも止水に入れると翅芽と尾鰓を大きく開く姿勢を取る.
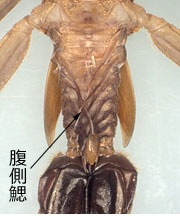
図14.ミナミカワトンボ科 コナカハグロトンボの腹側鰓.
Ⅱ.温度
(2-1) 耐性と馴化
幼虫の生活環境で「温度」というと,どれくらいの低温や高温に耐えられるかといった点がまず思い浮かびます.つまり高温・低温の耐性温度という視点です.しかしこれを実験的に厳密に調べてもあまり生態学的な意味はないことが多いと思われます.というのは,馴化 (じゅんか) という,一種の慣れによって,徐々に上下する高温・低温には意外と耐えてしまうことが知られているからです.ですから実験するにしても,種ごとに比較できる実験条件の標準化が難しいでしょう.そして自然界では,日単位,年単位で見ても,急激に温度が変化することは少なく,徐々に上がったり下がったりしますから,馴化の問題を見過ごすことができません.
この馴化のしくみですが,温泉が流れ込む川で,15~40℃の範囲に生息する北米産イトトンボ
Argia vivida 個体群内の酵素の変異について調べられた研究があります.これによると,グルコース6リン酸脱水素酵素をはじめとした4種類の重要な酵素について,アイソザイム
*3を発現して立体構造を変化させ高温でも活性を失わないようにしている,という馴化のしくみが明らかにされています (Schott & Brusven, 1980).
ところで,自然界をグローバルな視点で眺めると,トンボ各種の分布,特に緯度方向の分布や垂直分布が温度によって影響を受けているという見方に反対する人はいないでしょう.したがってトンボと温度の関係を考える際に,温度の絶対値より,それと相関性のある緯度や標高との関係で考えていく方が,理解がしやすいと思います.
ただこの場合,各地域の温度が,成虫,幼虫,卵,いずれに効いて分布が決まっているかははっきりしません.しかし一般論として,成虫はその地域でもっとも暖かいシーズンに出現するものであって,移動力があり,温度変化に対しては行動で避けることができるステージです.対して多くのトンボの幼虫は冬を越すステージであって,低温にいかに耐えるかが重要な生存戦略となります.もちろん成虫および卵越冬するトンボについては,それらのステージの越冬戦略が重要になります.以下はそういった意味で,主に幼虫に関わる問題として考えていますが,一部は成虫や卵の問題でもあります.
(2-2) 標高
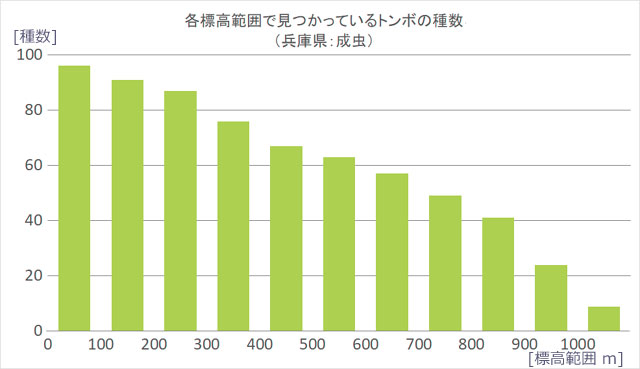
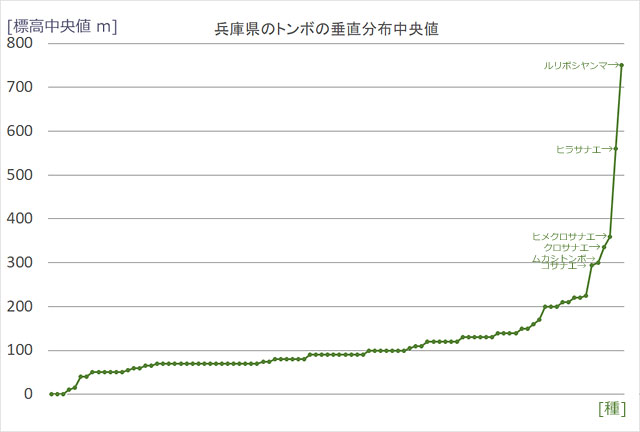
ではまず標高について,兵庫県に分布するトンボを使って分布状況をみてみましょう.兵庫県でもっとも高い山は氷ノ山で1,510mですが,だいたい1,000mを越えるあたりまでの分布調査結果をもとに,100m刻みで各標高範囲に何種類ぐらいのトンボが見つかっているか調べてみました (図15).また,飛来種を含めた98種について,見つかっている場所の標高分布の中央値で並べてみました (図16).
図15.兵庫県において各標高範囲に分布するトンボの種数.
標高が高くなるほどトンボの種数は少なくなっている.これは高標高地の低温が低地のトンボの進出を抑制しているためかも知れない.ただし,標高の高いところは生息地が少ないことや,ほとんどが山間の池や川という提供環境の多様性が少ないことなども考慮する必要はある.
図16.兵庫県の98種のトンボを分布の中央値の低い順に並べたグラフ (東・青木,2025参照).
兵庫県ではおおよそ200m以上が山地になる.標高の高いところに生息する2種およびコサナエは,いずれも北方に分布中心がある種である.ヒメクロサナエ,クロサナエ,ムカシトンボは山地の源流域に生息する種である.
図15を見ると,各標高範囲に分布するトンボの種数が.標高が高くなるほど少なくなっているのが分かります.これは,多くの平地のトンボが,気温の低い高い山に進出できないことをある程度反映した結果だと思われます.また標高の高いところに分布する,ルリボシヤンマとヒラサナエ (モイワサナエ中部個体群) の2種は,気温の低い北方に分布中心がある種です.兵庫県では,これらの種は気温の低い高標高地に見られる (歴史的に残っている遺存種) と,解釈できます.
このように,標高差による温度差が,トンボの分布に影響を与えていると考えられます.これらは誰もが思いつく当たり前のようなことですが,実際にデータで押さえておくことは重要です.同じことを,3,000m級の山がある,もう少し広い地域で行うともっとよい結果が出るかも知れません.
(2-3) 緯度
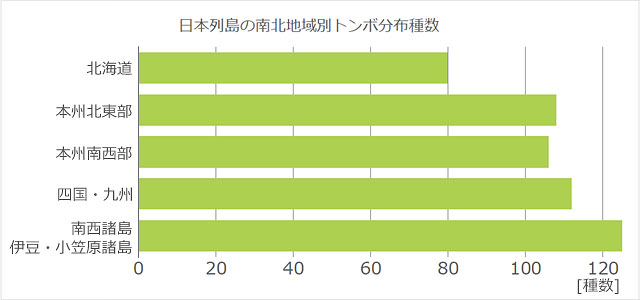
次に緯度について考えてみましょう.これも身近な例で見てみましょう.日本列島は南北に長く,北緯20度 (沖ノ鳥島) から46度 (択捉島) の範囲にあります.緯度で区切って分布を調べるのは難しいので,ここではおおざっぱに,北海道,本州北東部,本州南西部,四国・九州,南西諸島・伊豆・小笠原諸島などという5地域に分けて,それぞれに分布するトンボの種数を調べてみました (図17).
図17.日本列島を5つの地域に分けてそれぞれの地域に分布する種+亜種数をグラフ化したもの.本州北東部と南西部の境界は石川県-茨城県のラインあたりだが,全国的な分布状況を見てどちらに所属するかを筆者が判断した.飛来種は除いている.分布は尾園ら (2022) を参考に描いた.南の地域の方が種数が多くなっていて,北の方に行くにつれ種数が減少している.
これを見ると,北の地域へ行くほど分布する種+亜種数が減少していることが分かります.南西諸島などの島嶼地域は,総合すると面積は小さいのに,種数が北海道に分布する種の1.5倍ほど多くなっています.この現象は,高校生物の教科書に載るほど多くの生物に対して成り立っており,例えば「熱帯など低緯度地方では種数が非常に多く,高緯度になるにつれて種数が徐々に減少する傾向は,多くの生物で共通してみられる (浅島他,2020)」とまとめられています.
トンボに話を戻すと,これから推察されることは,熱帯地方で進化したとされるトンボが北方へ分布を広げるためには,さまざまな方法で低温に耐えるしくみを進化させる必要があり,これはそれなりに困難をともなうのだろうということです.そうであれば成功者が減り種数がだんだんと減っていくことになります.
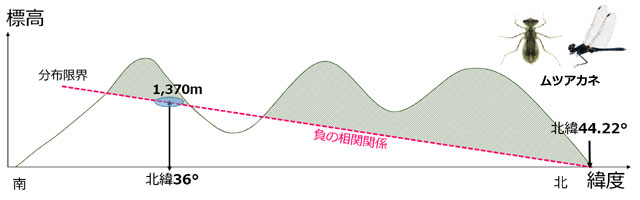
以上,標高と緯度に関することをまとめると,温暖地のトンボは南に行くほどより高地に生息でき,寒冷地のトンボは北へ行くほどより低地に生息することが予想されます.つまり大雑把に言って,分布限界は緯度と標高に関して負の相関関係を示すといえるでしょう (図18,Corbet, 1999).
図18.標高や緯度の違いによる分布の制限が温度によるものと考えたとき,分布限界は負の相関関係となるであろう.
ムツアカネは,実際は北緯36度では標高がもっと低いところに生息が可能かも知れないが,この南限生息地 (北緯36度,標高1,370m) を標高・緯度における分布下限と仮定すると,平地に生息できるのは北緯44.22度となる.
例として,寒冷地のトンボであるムツアカネで試してみましょう (図18).ムツアカネの国内の南限分布地は北緯36度の標高1,370mの地点です.気温減率を0.6℃/100mとすると,この地点は標高0m地点より8.22℃低いことになります.一方日本では緯度1度で気温が1℃ほど変化することが知られています.計算すると,この地点から北へ緯度にして8.22度行った地点,北緯44.22度における標高0mの気温が,このムツアカネの南限生息地と同じになります.これは北海道内のやや北の方になります.
とてもラフな予想ですが,北海道では平地に生息していることになるでしょう.そこで図鑑で確かめてみると,「北海道では平地や丘陵地...に生息するが,...本州では標高の高い山地の池沼に限られる (杉村ら,1999)」,また北海道のトンボ図鑑では「平地から山地の池沼,高層湿原に生息する (広瀬ら,2007)」などと記されており,この計算結果がだいたい当たっていることが分かります.
以上の検討から,トンボの分布には温度が大きく影響しているといえそうです.
最後に少し極端な例をお話ししておきましょう.兵庫県では寒冷地のルリボシヤンマが,標高の高い場所に生息していることを述べました.これと逆の例がないか探してみたところ,それらしいのがありました.オオシオカラトンボです.この種
Orthetrum melania は国内では南西諸島から北海道に至るまで分布していますが,「北海道では産地が限られて珍しく... (杉村ら,1999)」と書かれており,どちらかというと暖かい地方の種です.そこで北海道のトンボ図鑑を見てみると,「北海道全域に記録が散在するが,温泉地に多い (広瀬ら,2007)」と記されていました.温泉地の水温はそうでない地域より高くなると予想されますので,これらは暖かいところを好む種が分布北限の近くでは水温の高いところに生息しているということであり,温度とトンボの分布に関して面白い事例を記述していると思います.
Ⅲ.水質に関するその他の要素
溶存酸素や温度は,幼虫にとっては生死に直結する問題を引き起こす要因といえますが,これ以外にも水質に影響を与える要素はいくつかあります.例えば塩分濃度,水質汚染,薬剤などです.しかしこれらはいずれもある地域に特化した問題といえるでしょう.ここでは塩分濃度と有機化合物による水質汚染について取り上げます.
(3-1) 塩分濃度
淡水といえども,水の中にはさまざまな塩分,つまり電解質が溶け込んでいます.電解質の濃度がトンボ幼虫の生存に影響を与えるとすれば,それは次の3点が中心になるでしょう.それらは,(1) 電解質そのものの毒性,(2) pH (酸性・アルカリ性)の問題,(3) 浸透圧の問題です.浸透圧は電解質以外の溶質についても生じますが,自然界の淡水に溶け込んでいる非電解質の多くは有機化合物で,これは水質汚染の項で扱います.
電解質そのものの毒性としては,特に重金属イオンが毒性の高いものとして問題になることが多いようです.多くは人体に対する毒性が調べられており,水銀,カドミウム,鉛などが環境問題の原因物質として指摘されてきた過去がありました.トンボにこれらがどのような影響を及ぼすかについての知見はあまり見つかりませんが,スズが羽化の失敗を招く (Jones, 1985) などという報告が散見されます.これら重金属がトンボに与える影響というのは,ある意味特殊な状況下での話ですので,研究対象となりにくいのかもしれません.
*3.アイソザイムとは,同じ化学反応を触媒するが,アミノ酸配列などが異なる酵素のことをいう.この場合だと,立体構造が変化して高温による失活が生じにくくなるという意味にとらえるとよい.
淡水のpHは,通常それほど極端な変動が起きません.これは水中には空気中の二酸化炭素が溶け込んで,これがpHの急激な変化を防ぐ緩衝物質としてはたらくからです*4.多くはpHは6~8に保たれています*5.
しかし実際の幼虫の生息場所の水域というのを考えると,そこは複雑な生態系ができており,生息している動植物の排せつ物や遺体が沈殿し分解されている環境です.一般にこういったものの分解産物として腐植酸が発生し,水域が酸性化することが多いようです (腐植酸性質).またコンクリートが多用されているため池などでは,水酸化カルシウムが溶け出し,大きくアルカリ性に傾くことが知られています.
また水域のpHの変化は,例えば金属イオンの存在様式を変化させたり*6と,さまざまな物質の存在様式に影響を与えています.したがって,pHのトンボ幼虫の生存に対する影響に関しては,複雑な生理化学的な議論が必要で,本サイトの範囲を超えそうですのでここでは割愛します.そこで,Corbet (1999) がいくつかの研究を引用して紹介している,カオジロトンボの話をしておきましょう.
*4.炭酸 重炭酸バッファと呼ぶ.
*5.pH=7が中性,それより値が低くなると酸性,高くなるとアルカリ性である.pH6~8というのは中性付近ということ.
*6.例えば,アルミニウムイオンは,酸性条件下ではAl+として,中性前後ではAl(OH)3として,アルカリ性では錯イオン[Al(OH)4]-として存在する.
カオジロトンボは北方性のトンボで,高層湿原に生息しています (欄外図19).高層湿原は腐植酸性質の水で,一般に酸性です.この例はスウェーデンでの話ですが,湖が汚染によって酸性化し,最上位の捕食魚が減少してきている状態でした.カオジロトンボは,魚の棲む酸性化していない湖より,そのような酸性化した湖によりふつうに見られました.その理由として,カオジロトンボ幼虫はあまり隠れようとせず捕食されやすい行動をとることを挙げています.つまり,酸性化したので捕食魚が減少し,捕食されやすい行動をとるカオジロトンボが多く生き残った,という論理です.
この例で分かるように,pHが直接トンボ幼虫に影響を与えるというよりは,pHが他の生物にまず影響を与え,その結果としてトンボ幼虫に影響が及んでくるという構造になっています.いわば一種の間接効果です.pHに限らず,水質のトンボ幼虫に対する影響は,このような間接的なことをも十分に考慮する必要がありそうです.

図19.カオジロトンボ幼虫.
最後に浸透圧の問題です.濃度の異なる水溶液を,水分子だけが通過でき溶質が通過できないような膜 (半透膜) で仕切ると,水が濃度の高い方から低い方へと移動します
*7.この水の移動圧力を浸透圧といいます.生物の細胞膜にはアクアポリンというチャネルがあってそこを水分子だけが通過できるので,半透膜の性質を有することになります.したがって,細胞内液 (濃度約0.9%) より濃度の低い水溶液 (もちろん濃度0の真水も含みます) 中では水が細胞内に進入し,逆に細胞内液より濃度の高い水溶液中 (たとえば海水,濃度約3.5%) 中では細胞内から水が外へ出て行きます.したがって,何らかの濃度調節機構が存在しなければ,体液の濃度が変化して生物は水中で生きていくことができません.ただトンボ幼虫に関していえば,海水中に生息するものはいませんので,問題は淡水中または汽水中の浸透圧調節ということになります.
トンボ幼虫の体表面はクチクラで被われ,通常体表から水が細胞内に進入してくることはありません.問題は呼吸器官です.これは水中から直接酸素を取り込む器官ですので,細胞が直接水と触れることになります.したがってここから細胞内に水が進入してくることになります.不均翅亜目の鰓では,淡水魚と同じようなしくみで,水が進入しても体液の塩分濃度が変わらないように調節しています.
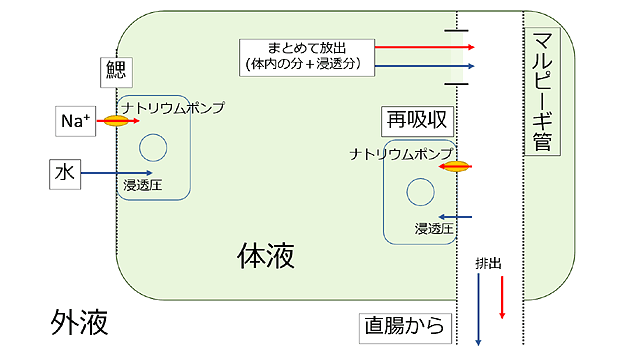
図22.不均翅亜目幼虫における浸透圧 (塩分濃度) の調節.
鰓では浸透圧によって水が進入してくる.同時にナトリウムポンプで外液からナトリウムイオン (塩分) を取り込む.これらはその後体液に移動し,体液中のナトリウムイオンや水と混じる.この混じった体液はマルピーギ管でいったん管内に放出される.マルピーギ管内ではナトリウムポンプによって体液に必要な少量のナトリウムイオン (塩分) が取り込まれ濃度が高まるので浸透圧が生じる.その浸透圧に応じてマルピーギ管から水も細胞内に入る.そこで再吸収されなかったものは排出される.この図には老廃物やその他のイオンなどは書き込んでいない.なお基本的しくみはヒトの腎臓も同じである.
水が進入する鰓には塩類吸収上皮があり,その細胞膜にナトリウムポンプというナトリウムイオンを取り込む装置が存在しており,これで淡水中の非常に低い濃度のナトリウムイオンを,より濃度の高い細胞内に取り込みます.濃度差があるのでこれにはエネルギーが必要で (能動輸送),塩類吸収上皮細胞にはそれを供給するためのミトコンドリアがたくさん存在します.
吸収されたナトリウムイオンと浸透圧によって進入してきた水は,老廃物やその他の物質とともに,マルピーギ管 (欄外図21) という器官で体液から管内に放出されます.ここでは進入してきた以上の水やナトリウムイオンが放出されるため,体に必要な水やナトリウムイオンを再吸収しなければなりません.ここのしくみにもナトリウムポンプがが活躍しています.マルピーギ管の中から細胞内へナトリウムイオンを能動輸送して細胞内の濃度を高めることで浸透圧が高まり,マルピーギ管内の水を,体に必要な分だけ細胞内へ再吸収することができます.そして余分な水と老廃物などは残されたナトリウムイオンとともに,回腸・直腸を通って体外に放出されます.
*7.図20.浸透圧の説明図.
半透膜とは溶質の分子は通さないが,水分子は通すというような性質を持つ膜である.細胞膜にはアクアポリンという水分子だけを通す孔がある.
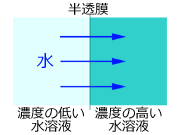
一見すると,濃度の低い方から高い方へ移動するという現象に違和感を覚えるかも知れない.これについては移動するのが「水」であるという点に注目すればよい.濃度の高い水溶液と低い水溶液を同体積とると,濃度の低い水溶液の方が溶質の分子数が少ないから,反対に考えれば水分子の数は濃度の低い方が多いことになる.したがって,水は多い方から少ない方へ「自然に」移動するのである.これを拡散といい,熱力学のエントロピー増大の法則に従っている.なお,浸透圧を扱うときの濃度は%濃度ではなくオスモル濃度を用いる.
図21.マルピーギ管の位置.
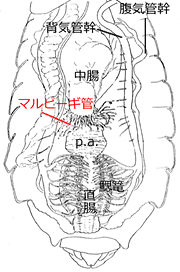
中腸とそれに続く嚢部 (prerectal amphulla) の接続部に糸状に広がっている.なお,これは腸管を見せるために切って短くして描いており,実際はもっと長い.ムカシトンボを背中側から見ている.Asahina (1954) を改変.
以上のように,一般にトンボ幼虫は濃度の低い外液中での浸透圧調節を行うしくみを発達させていますが,濃度の高い外液中では浸透圧調節ができません.それは体液の約3倍の塩分濃度 (約3.5%,1,000オスモル) を持つ海水中にトンボ幼虫がいないことと関係がありそうです.
これに関して,実験的にどれくらいの外液濃度まで調節ができるかを調べた研究者がいます (欄外図23).Aeshna cyanea というルリボシヤンマ属幼虫を使って,外液濃度を変化させたときの体液の濃度を調べています.外液の濃度を300オスモルぐらいまで上げても,体液の濃度はあまり変化していません.これは調節が効いている証拠です.しかしそれより外液の濃度を上げると体液の濃度も上昇してしまっています.つまり調節ができなくなってしまったということです.300オスモルという濃度はだいたい体液の濃度に一致しますので,体液と同じ濃度くらいまでは調節可能ということになりましょうか.
この調節能力は種ごとに異なっているようで (Corbet, 1999),汽水域に棲むヒヌマイトトンボなどではもう少し高い調節能力を持っているかもしれません.
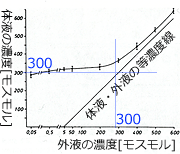
図23.外液の濃度を変化させたときの体液の濃度変化.
Aeshna cyanea の F-1齢幼虫を使っている.縦線は10個体の標準偏差.Komnick (1978) を改変.
(3-2) 有機化合物による水質汚染
水質を汚染する物質は多岐にわたります.トンボにとって最近よく話題になるのは,ネオニコチノイド系殺虫剤です.また1960年代前後から,有機塩素系殺虫剤 (DDT,BHCなど),有機リン系殺虫剤 (マラチオンなど),その他の殺虫剤,重金属などによる汚染が問題にされ,公害問題と結びつけて語られました.これら殺虫剤の問題は重要ではありますが,ヒトとトンボの関わりの中で生じている問題であって,自然の中で進化してきたトンボ幼虫にとっては,本来出会うことのなかったものといってよいでしょう.トンボとはどういう生き物かを知るという観点に立つと,この問題は別の文脈で扱った方がよいと考え,ここでは扱わないこととします.
そこで,自然にも大なり小なり起こりうる問題として,有機化合物の問題についてみておくことにします.公害問題が生じていたころは,家庭排水,し尿,また製紙排水などが大量に水系に流され,それに含まれる有機化合物が原因する富栄養化が問題にされました.ただ有機化合物の水域への流入は自然にも起きており,むしろ適度な富栄養化は生態系を支える生産力につながるものだという重要性が,現在ははっきりと認識されています.
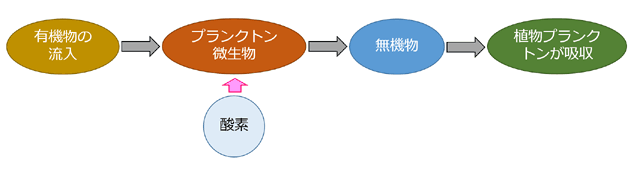
図24.水域に流れ込む有機化合物の分解過程(水質浄化)の模式図.
水域に流れ込む有機化合物は,動物プランクトンその他の動物および微生物など,従属栄養生物にとって栄養分となる,それらを代謝分解するのは基本的に呼吸作用であるので,水域の酸素を消費する.食物連鎖を通じて最終的に排出された無機化合物は植物プランクトンや藻類などの植物 (独立栄養生物) の養分として消費される.この時点で水域に流れ込んだ有機化合物は水の中から除去されたことになる.
富栄養化とは,図24で,水域中の無機化合物濃度が高まることを意味しています.富栄養化すれば,植物が増え,それを出発点とする食物連鎖が太いものになって,魚介類などの豊かな幸がいっぱい増えることになります.生物が豊かな生態系を支える基盤になるのがこの無機化合物,つまり富栄養化です.
しかし有機化合物の流入が大きすぎると,それを餌とする動物プランクトンが増え,さらに植物プランクトンも増えていきます.これらはやがて死滅し大量の死骸 (有機化合物) となります.こうやって有機化合物が再生産されて雪だるま的に増えていきます.これら有機化合物は水質浄化の流れに入ることになり,そのときに大量の酸素を消費します.その結果,水域中の酸素が不足したり時には無酸素状態になったりして,水域の魚介類が死滅します.これが「行きすぎた」富栄養化によって起きる問題です.淡水域で生じる現象としては,藍藻類が主体となったアオコの発生です.
なお,自然のバランスが保たれているところでは,流入および再生産される有機化合物の量が少なくて,水質浄化が行われても酸素が不足するまでには至りません.
水質浄化が行われる場所は,水中よりも,有機化合物が堆積する水底です.トンボ幼虫の多くは水底で暮らしています.したがってここで溶存酸素量の問題が生じることになります.底生幼虫の形態や行動に関しては,すでにお話ししました.またこれはトンボ幼虫に対してだけ影響を与えるものではなく,餌生物にも影響を与え,間接的にトンボ幼虫に影響を与える場合もあります.
トンボの生態学 5.幼虫の生活
他の生物との関わり-1.捕食者
前節で検討した温度や溶存酸素量は幼虫の生死に関わる問題ですが,捕食者や餌生物などまわりの生物との関係性も,幼虫の生存にとって大きな圧力になります.ここではこういった生物的環境について見ておきましょう.
トンボ幼虫の捕食者といえば,まず頭に浮かぶのは昆虫食の魚類でしょう.またトンボ幼虫に接している方なら,例えば小さな均翅亜目の幼虫はヤンマのような大型の幼虫に捕食されることもよく目にされていると思います.さらにまた捕食者を広く定義するならば,寄生者や病原体なども捕食者といえるかもしれません.ここでは昆虫食魚類と大型の捕食性昆虫について考えますが,その前に寄生者について簡単にふれておきましょう.内部寄生虫は幼虫から成虫へと連続して寄生することが考えられますので,以下はそういった点にも注意してください.
Ⅰ.寄生者
Pavlyuk (1978) は,ウクライナ西部で,全58種におよぶ幼虫500頭,成虫16,046頭のトンボから,87,140個体の寄生虫を得ています.そのうちもっとも多い52.61%が原生動物門・胞子虫亜門・グレガリナ亜綱に属するもので,続いて32.87%が節足動物門・クモ形綱・ダニ目に属するいわゆるミズダニ類の幼虫,14.32%が扁形動物門・吸虫綱に属する寄生虫のメタセルカリアであって,これで全体の99.8%を占めています.あと少数のものは,扁形動物門・条虫綱が0.13%,袋形動物門・線虫綱が0.06%,ということです (四捨五入により100%にならない.以上は五界説による分類).このうちミズダニ類については成虫の寄生者ですので,ここでは扱いません.
もっとも多いグレガリナ類はトンボが終宿主になっているようです.一般的な生活環は,餌昆虫に付着した嚢胞体を摂食したときに感染,腸管から体内へ侵入し無性的に分裂増殖したあと,有性期に入って接合子を形成し糞とともに体外に排泄されるというものです.そしてこの接合子から嚢胞体が形成され餌生物に寄生することで生活環が完了します.成虫への寄生がよく確認されるようです (Åbro, 1974).
吸虫類というと,日本吸血住虫というヒトへの寄生虫がよく知られています.セルカリアという幼虫のようなステージで皮膚から侵入してヒトに感染します.トンボの場合,幼虫が第一中間宿主である巻き貝を食することによってセルカリアが侵入し,体内で次のステージであるメタセルカリアになります.トンボは第二中間宿主で終宿主のカエルや鳥などの脊椎動物に食べられることで成虫となります*8,
最後に,Pavlyuk (1978) の報告ではわずか0.06%でしかありませんでしたが,袋形動物門・線虫類に属するハリガネムシについて考えておきましょう*9.ハリガネムシはカマキリの肛門から出てくることでよく知られています.しかし元来は淡水中で生活しているもので,非常に小さな卵がデトリタス・フィーダーに食べられて,その体内に寄生するところから寄生生活が始まります.寄生するとシストを形成して休眠します.カマキリなど直翅目は,これらデトリタス・フィーダーの成虫 (ユスリカ,カゲロウ,カワゲラ,トビケラなど) を捕食することで寄生されることになります.そしてこれら宿主を水辺へ誘導して水に飛び込ませる行動を誘発し,成虫が水中へ戻るそうです (Sato et al., 2011).
トンボに関しては,幼虫では見たことがありませんが,成虫でハリガネムシが肛門から伸びているのを見ています (図26).トンボ幼虫はユスリカなどデトリタス・フィーダーの捕食者ですから,幼虫時代に寄生される可能性があります.一方成虫もカマキリと同じでこれらデトリタス・フィーダーの成虫を捕食しますから (図25),その経路で寄生される可能性もあります.いずれにしても水辺や水中で生活するステージを持つトンボは,ハリガネムシにとっては都合よい宿主になるでしょう.
以上寄生虫に関しては,筆者は門外漢であり,興味のある方はいろいろと調べてみてください.
*8.吸虫類の基本的生活環
終宿主(カエルなど脊椎動物)
↓
卵が排泄物に混じって体外に放出される
↓
孵化してミラシジウムとなり,繊毛を持っており活発に泳ぎ,巻き貝に侵入
↓
第一中間宿主(巻き貝)
体内でスポロシストとなり,レジアを消化腺の中に放出すると,レジアはセルカリアとなる
↓(採餌)
第二中間宿主(トンボ幼虫)
体内でセルカリアはメタセルカリアとなる.トンボ幼虫は成虫となることもある
↓(捕食)
終宿主(カエルなど脊椎動物)
メタセルカリアは成虫となる
*9.現在は類線形動物門・ハリガネムシ綱に分類されている(高知大学ホームページ).
https://www.kochi-u.ac.jp/
w3museum/Fish_Labo/
Member/Endoh/Metazoa/
nematomorpha.html

図25.トビケラ類を捕食するヒラサナエのオス.
図26.ハリガネムシに寄生されたトンボたち.左はヨツボシトンボのメス,右はアオモンイトトンボのオス.
Ⅱ.捕食者との関わり
トンボ幼虫を捕食する生物としては,昆虫食魚類,水生の鳥類,トンボ幼虫などが思い浮かびますが,爬虫類,両生類も捕食者となっている例が報告されています.こういった捕食者の中で,最近は,例えばオオクチバス (欄外図27) などがよく引き合いに出されます.これは重要な環境問題ではありますが,日本のトンボたちは進化の過程で長い間この外来魚に出会ったことがありません.トンボたちがいかに進化して現在の姿になったか,すなわち適応*10を考える際には,こういった最近の外来生物の問題は,むしろノイズになる可能性があります.したがってこれらの問題はまた別にところで考察することとして,ここでは扱いません.
(2-1) 隠蔽 (隠れること)
昔からその生息地で共存している捕食者に対して,トンボ幼虫はさまざまな方法で対抗する手段を進化させています.まず最初に思いつくのは擬態でしょう.擬態のうち隠蔽的擬態とは,幼虫がその生活場所の背景に溶け込むような色彩や形態をしていて,他の動物から見つかりにくくなっているような状態をいいます.トンボ幼虫は魚類などの被食者であると同時に,餌動物の捕食者でもあるので,幼虫の擬態には餌に近づくためという側面もあることを忘れてはいけません.

図27.捕食者としてよく引き合いに出されるオオクチバス.産卵床のまわりをゆっくりと泳ぐ.
*10.適応とは,進化の結果身についた行動や形態等を指す言葉である.いわば遺伝子とその発現の過程にその形質が組み込まれている.対して個体が生まれてから身につけた行動や形態等は,学習,馴化 (慣れ),などという用語が使われる.
隠蔽的擬態の有様は,色や斑紋に現れることが多いと思われます.そこでトンボ幼虫の色や斑紋について少しふれておきましょう.幼虫の泥を落とすと,そこには特徴的な色や斑紋が姿を現します.このうち,基本的には,色 (地色) は真皮の色で,斑紋は表皮 (クチクラ) が暗化した部分です
*11.これは幼虫の斑紋と羽化殻の斑紋を比べると分かります.羽化殻はクチクラが真皮から離れて残されたものなので,これに黒い斑紋が残っていることから斑紋はクチクラに付いた色であることが分かります(図29).
図29.ヒナヤマトンボの幼虫と羽化殻の斑紋.
幼虫には,黄色の地色中に濃褐色の斑紋がある.羽化殻を見ると,黄色の地色の部分は透けており斑紋の部分は残っている.つまり黄色の地色は真皮の色であり,斑紋はクチクラの色であることが分かる.
幼虫の全体的な色や模様は,この地色と斑紋が混じって形成されています.地色は単色であるとは限らないようですし,クチクラにも全体的に色が付いていることがあります.また筆者の経験から,体色は脱皮時のまわりの環境の影響を受け濃くなったりうすくなったり,色が変わってきたりしますが,斑紋パターンはその影響を受けずに種特異的に出てくるように思います(図30).
*11.昆虫の皮膚の構造.
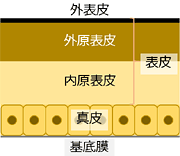
図28.昆虫の皮膚の一般的構造は,大きく分けて表皮・真皮・基底膜からなる.表皮 (クチクラ) は,外表皮 (数層からなる) と原表皮 (外原表皮+内原表皮) からなっている.真皮は生きた細胞からなっており,基底膜は体腔と皮膚を隔てている.図および用語は後閑 (1984)を参照した.
図30.ヒメサナエの地色と斑紋.
この2個体のヒメサナエ幼虫は大きく地色が異なっているが,斑紋のパターンは両者でほぼ同じである.この地色の違いが,クチクラの色なのか真皮の色なのかは不明である.
図30のような地色の違いはしばしば見られます,ムカシトンボでは黄褐色の幼虫と黒褐色の幼虫が見られます (図31).このような体色の違いは生息場所の背景色を反映していると見られ,脱皮直後の体色の発現時に背景に合わせた色になるようです.飼育例として,図32にオキナワミナミヤンマ幼虫を示します.背中が灰褐色をした亜終齢幼虫を茶色の落ち葉を入れた容器で飼育していたとき,脱皮した終齢幼虫が全身黒褐色になりました.現地で得た終齢幼虫はそれほど真っ黒ではありません (図32).

図31.体色の異なるムカシトンボ幼虫.

図32.オキナワミナミヤンマの体色の変化.
褐色の落ち葉を敷いた飼育容器で脱皮したオキナワミナミヤンマの幼虫は体色がほぼ真っ黒になって,現地で採集されたものとは体色が異なる.なお腹部は本種の場合もともと黒褐色をしている.
さてトンボ幼虫で,我々が気づきやすい隠蔽的擬態の例としては,ギンヤンマやムカシトンボの若齢幼虫に現れる白黒模様があります (欄外図33).これは,光と影が混じり合う水面近くで生活する幼虫の体をとぎれとぎれに見せ,その正体をわかりにくくする効果があると言われています (Corbet, 1999).またアオハダトンボの細長い幼虫はツルヨシなどの植物沈積物の中に紛れ込むとその中の枯れ茎のように見えます (図34左).砂地に浅く潜って生活するキイロヤマトンボのゴマ塩模様は砂粒に同化し,捕食者に見つかりにくくなると同時に,近づく餌に気づかれにくいという側面もあるように思えます (図34右).
ただこれらは,我々人間の感覚によってそのように感じているだけで,トンボの視覚はヒトと異なりますので,これらの擬態が実際に効果があるかどうかは,実験を計画して確かめないといけないことは指摘しておきたいと思います.

図33.白黒模様のムカシトンボ若齢幼虫.
図34.アオハダトンボの幼虫とキイロヤマトンボの幼虫の隠蔽的擬態.
左はアオハダトンボ幼虫で,ツルヨシの枯死体につかまり,流れに身を任せて植物沈積物の一部のように擬態している (野外観察).右はキイロヤマトンボ幼虫が砂地に潜っているところで,背中や脚にあるごま塩模様がまわりの砂粒にうまく溶け込んでいる (飼育観察).
さて,底生生活をするトンボ幼虫の中には,底の砂泥に身を埋めて生活する種が結構います.特にサナエトンボ科やオニヤンマ科に多いのですが,これらの幼虫は通常泥を身につけてカモフラージュしています.これらは体色や斑紋を進化させて擬態しているのではなく,隠蔽工作を行って生活している幼虫たちといえるでしょう.厳密な定義ではこれは擬態とはいえないかもしれませんが,ここでは定義を拡大して,これも一種の隠蔽的擬態としておきたいと思います.
図35から分かるように,ヤマサナエの幼虫は,体色や斑紋 (c) が,(a) のような砂泥底の背景の中に紛れ込めるように進化しているわけではありません.砂泥底で長い間生活を続けてきた結果,細かい毛を体表面に密生させ (e),自然に泥が体表面に付着するような形質 (b) を進化させてきたと考えるべきでしょう.幼虫を採集した直後には,うすく体表に付着したシルトのような泥は落ちません (b).また羽化殻が常にうすく泥をかぶっている (d) ことからも,自然に落ちるようなことはないことが分かります.つまりこれは,一つの形質 (毛) が,その生活場所が有する特質 (泥) から必然的に隠蔽的な外観を誘導すること (泥をまとう) で発現する,二次的な擬態であると言ってよいでしょう.
図35.ヤマサナエは泥を身につけて水底に隠蔽する.
(a) 実際の川底に潜っている幼虫.(b) (a) の状態の幼虫を採集し軽く水でゆすいだもの.(c) (b) の状態の幼虫を歯ブラシで洗浄し泥を完全に取り除いたもの.(d) ヤマサナエの羽化殻は通常泥で被われた状態で発見される.(e) 体表面に細かい毛が密生している.
同じ状況は,ヤマサナエの属するアジアサナエ属,ホンサナエ属,ミヤマサナエ属など砂泥底に浅く潜って生活する幼虫に当てはまります.一方で,砂礫底で生活するアオサナエ (欄外図36),オナガサナエ,オジロサナエ,ヒメサナエなどの幼虫は,多少の泥がついている場合もありますが,細かい毛が生えてシルト状の泥が背面全体に付着するようなことはありません.同じ底生幼虫でも,生活 (進化) してきた環境の違いがこのような形質の違いになって現れているといえるでしょう.
(2-2) 逃避と防御
トンボ幼虫は,捕食者から隠れること以外に,いくつかの形質を進化させて,捕食者に対抗しています.まずは捕食者からの逃避です.捕食者に見つかったと自覚した場合,直接の反応としてその場を離れようとします.
不均翅亜目幼虫の場合,アオサナエのように,肛門から水を吹きだして「ジェット推進」できる種がいます.アオイトトンボでは体をくねらせて泳いだり,脚で水をかいて直線的に泳いだりします.ただしこの逃避方法は,捕食者が魚の場合,おそらく成功しないと思われます.むしろ魚の注意を集めてしまい,あっという間に捕食されてしまうように思います.ですから,この方法が有効なのは,魚ほど泳ぎがうまくない,ヤンマなどの大型トンボ幼虫をはじめ,ゲンゴロウ幼虫,タガメなど,昆虫の捕食者の場合だけだろうと思います.
また幼虫が捕まってしまうなど,捕食者から逃げられないと悟ったとき,反撃的な行動に出ることがあります.下唇で噛みついたり,尾部付属器で突き刺したりする攻撃行動です.これはヤンマなどの大型幼虫を手づかみしたときに,噛みつかれたり,腹部を曲げて尾部付属器で突かれたりする経験からも,想像できます.捕食者と体格差があまりない場合には有効な方法かも知れませんが,大型の捕食者の前では役に立たないかも知れません.しかし最後の手段としての生存をかけた戦いという位置づけは可能でしょう.これらの攻撃行動がどの程度トンボ幼虫の生存率を高めているかについては,まだ推測の域を出ていないようです.
これらの直接的な逃避・攻撃以外に,もう一つ別の手段を持っている種がいます.クワガタの採集方法の一つに「木を蹴ると落ちてくるのでそれを拾う」というのがあります.私は子供のころ裏山でこの方法で大きなノコギリクワガタをたくさん捕まえたのを覚えています.強い振動を感じると,クワガタは脚を側部に折り畳み硬直した状態になって,落下します.これを擬死または不動反射と呼びます.トンボ幼虫にも擬死をするものがいます.兵庫県のトンボでよく見るのはコシボソヤンマとムカシトンボです.

図36.アオサナエ幼虫(野外).
多少の泥はかぶっているが腹部背面など地色が見えている.
図37.コシボソヤンマとムカシトンボの擬死(不動反射).
左はコシボソヤンマの擬死で,どの個体でもこの反り返った同じ姿勢をとる.右はムカシトンボの擬死で,脚を両脇に密着して動かなくなる.
擬死に関しては,Miyatake et al. (2004) の論文が有名です.これは,コクヌストモドキという体長3~4mmの甲虫と,それを捕食するクモ,アダンソンハエトリ(体長はオス:5~7mm,メス:6~9mm)を使った実験的研究です.コクヌストモドキの飼育株から擬死継続時間の長い系統と短い系統を10世代にわたって人為選択し,11世代目の長い系統と短い系統をアダンソンハエトリに捕食させて,それぞれの成功率の違いを比較するという実験です.
その結果,擬死継続時間の長い系統の方が捕食にあう頻度が少ないことが示されました.さらにまた各系統の人為選択において,長系統では世代を追うごとに擬死継続時間が長くなっていくことも示されました.以上をまとめると,この2種の捕食-被食関係では,擬死継続時間の長い個体が選択され (生き残る確率が高くなり),擬死がより強く維持される方向に進化が進むということになります.すなわち擬死は捕食に対する適応である,ということです.
トンボ幼虫の擬死に関して,新井 (2001) は,擬死は捕まえられるなどの刺激を受けたときに生じるので,「捕食者にとらえられたあとで,あわてて死んだふりをしたところで助かる見込みはないように思う」と述べています.しかし魚がいったん餌を吐き出せば,そのときに起きる擬死が2回目の捕獲を起きにくくするかも知れません.
コシボソヤンマは少しの刺激で強い擬死反応を引き起こす (図37左) 流水性のトンボで,これはほぼすべての個体に見られます.この事実は擬死に対して強い選択圧がはたらいている証拠と見ることができます.またコシボソヤンマの体表には,たくさんの棘があります (欄外図38).コシボソヤンマが食われて魚の口の中に入ったとき,側棘や尾部付属器の先で魚の口内を攻撃すれば,あるいは攻撃しないまでも口内で擬死したらその姿勢 (図37左) から尾部付属器なとの棘が口内に突き刺ささって,魚は思わずこれを吐き出すでしょう.そのとき擬死しておれば,魚はトゲトゲの無生物を口に入れたと勘違いし,それを再び口に入れようとしないかも知れません.しかもこれは流水環境で起きますから,吐き出された擬死幼虫は物体と化して流れ去り,魚の視界から直ちに消えてしまうことになります.クワガタが落下して見つかりにくくなるのに類似した状況です.これは実験が可能な仮説で,調べてみる価値がありそうです.
新井 (1987) は擬死を行う日本産トンボ幼虫を調べており,属数にして15報告しています
*12.そのうち10属が流水性種で,3属が湿地性種です.流水性種が多く含まれていますが,これは流水環境 (つまり流されて視界から消えること) が擬死に有利にはたらくことを反映しているからかも知れません.
トンボ幼虫の擬死についてはまだまだ研究がなされていませんので,推論はこのあたりにしておきましょう.擬死に関しては,擬死そのものを一般的に扱うだけでは不十分で,どういう生態的条件の中で擬死が有利にはたらくかということを,種個別的に調べる必要があると考えています.
ところで,幼虫の棘と捕食魚の関係について調べた論文があります.カオジロトンボ幼虫には,腹部第9節側棘の長い個体と短い個体がいることが知られています (図39).これらは同じ水域の個体群内に混在しているわけではなく,一つの水域では同じタイプの個体が棲息しています.
図39.側棘の長さの異なるカオジロトンボ幼虫.腹部第9節の長さが肛側片より短いもの(左)と長いもの(右).かつては北海道産は長いといわれていたが,横山 (2016) の調査によって,北海道にはこれが短いものや,肛側片の長さと等長のものがいることが明らかにされた.
Johansson & Samuelsson (1994) は,スウェーデンで,魚の棲息している7つの止水域と魚のいない9つの止水域を選び,それぞれに棲息するカオジロトンボ終齢幼虫の棘の長さを測定しました.彼らは,腹部第9節側棘以外に,腹部第4,6節の背棘の長さも測定しました.その結果,魚の棲息する水域で採取された幼虫の各棘の長さは,魚のいない水域でのものより有意に長いことが示されました.前節でカオジロトンボはpHの低い池沼に棲息していると書きましたが,各棘の長さはpHとは無関係であったと述べています.また各棘の長さは体長との相関性は認められず,一方でこれら3つの棘の大きさはそれぞれ相関性があったと記しています.さらに,魚のいる水域の幼虫の個体数は,いない水域より有意に少なかった (つまり捕食圧が存在する?) ということです.彼らは,捕食魚ヨーロピアンパーチにそれぞれの幼虫を捕食させる実験も行っており,棘の長い個体の方の処理に有意に多くの時間を費やしたことを確かめています.
以上の結果から,捕食魚の存在が各棘の長い幼虫の生存率を高める (食べられるのに時間がかかるから) こととなり (選択圧),次第に棘の長い個体ばかりになっていった (小進化) と考えることができます.これは,棘の長さが幼虫の生存率を高めることを意味し,棘は身を守る防御機能を果たしていることが示唆されます.
横山 (2016) は,北海道の4つの水域を選んで,それぞれの個体群の幼虫の側棘の長さ (と腹部の斑紋) を調べています.また私は本州中部の高層湿原の池塘で側棘の短いカオジロトンボ幼虫を採取しています (図19).インターネットでそれらの水域の写真を比べてみますと,側棘の長い個体が棲息する場所はいずれも大きな水面が広がる水域を有し,短い個体が生息する場所は,湿原のあちこちに小さな池塘が点在するような水域でした (図40).池沼が大きければ魚がいる可能性が高く,これは Johansson & Samuelsson (1994) の結果に一致する可能性があるといえるでしょう.

図38.コシボソヤンマにはたくさんの棘があってとてもいかつい幼虫である.日本産ではアオサナエ幼虫にも棘がたくさんあり同様に擬死をする.
*12.擬死を行うトンボの属
(流)流水性種,(湿)湿地性種
アオヤンマ属(湿)
トビイロヤンマ属(湿)
コシボソヤンマ属(流)
サラサヤンマ属(湿)
ミルンヤンマ属(流)
オオヤマトンボ属
コヤマトンボ属(流)
ムカシトンボ属(流)
ヒメクロサナエ属(流)
アオサナエ属(流)
オナガサナエ属(流)
コオニヤンマ属(流)
ヒメサナエ属(流)
オジロサナエ属(流)
ヨツボシトンボ属
新井 (1987) による.
図40.カオジロトンボの棲息する水域景観の違い.
左は本州中部の高層湿原で小さな池塘が点在する水域.ここには棘の短いカオジロトンボ幼虫が棲息する (図19).右は北海道の低層湿原で大きな水域が広がる.ここには棘の長いカオジロトンボ幼虫が棲息する (図39右).
最後に均翅亜目の例を見ておきましょう.均翅亜目幼虫はいずれも弱々しく見え,他の捕食者の餌的存在であるように見えます.そんな均翅亜目が持っている逃避手段に自切というのがあります.いわゆるトカゲの尻尾切りと同じで,捕まったとき,尾鰓や脚を自分で切って逃避するという方法です.例えば尾鰓ならその付け根というように (欄外図41),自切できる部分は決まっており,自切した次の脱皮以降,それらの部分は再生されていきます.この現象は,均翅亜目幼虫を飼育しているときに,よく観察されることです.

図41.左尾鰓を自切したグンバイトンボ終齢幼虫.
トンボの生態学 5.幼虫の生活
他の生物との関わり-2.採餌
トンボは例外なく肉食です.したがって生きていくためには,餌となる他の動物を捕まえる必要があります.トンボの餌となる動物は,主に自身より小さな無脊椎動物ですが,ときにメダカのような小さな魚や両生類の幼生なども餌にすることがあると思われます.これら脊椎動物を捕食する場面は,野外ではなかなか目にすることはなく,飼育下での観察によって確かめられています.そして意外によく食べられているのは巻き貝だそうです.
トンボ幼虫が餌を捕る方法は,種ごとに異なっているといえますが,大きく分けて「待ち伏せ型」か「狩猟型」に分けることができます.状況によってはこれらを使い分ける種もあるようです.
(1-1) 餌を捕獲する下唇の形態と餌の捕らえ方
よく知られているように,トンボの餌捕獲の武器は伸縮する折り畳まれた下唇
*13です.これを筋肉の力や,肛門を閉じた上で横隔膜を収縮させ (図3b),圧力を前方に伝えて (呼吸では肛門を開けて水が肛門から吹き出る),伸ばし,餌を捕獲します.下唇をよく観察すると,実にいろいろな形態をしていることに驚かされます.これは各種の餌の捕り方や,生活場所で主に捕っている餌動物の種類との関係で,長い時間かけて進化してきた形態なのでしょう.
よく言われているのは,下唇前基節の形状を大きく分けて板状のものとスプーン状のものがあるというまとめ方です (図42).これは大きく言えばその通りですが,板状に見えるものでも,下唇側片が少し下唇前基節の平面に対してねじれて位置し浅いスプーン状に見えるものがあり,この区別はやや主観が入り込む余地があります (図44).
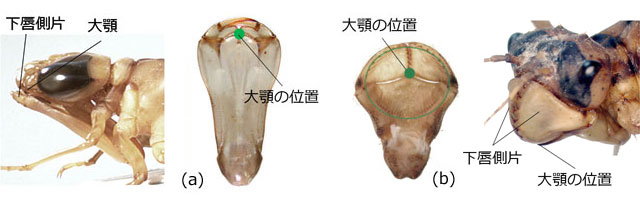
私は,餌の捕り方から考えて,可動鉤の長さおよび側刺毛・腮刺毛の発達程度が,重要なポイントになるのではないかと考えています.それらについて例示する前に下唇前基節の共通点を一つ指摘しておきましょう.トンボの口器には,左右に開く鋭い歯状の大顎と呼ばれる器官があって,これで餌生物を噛み砕きます.この大顎の当たる位置がおおよそ下唇中片の先端部付近になるのです (図42).ですから捕らえた餌をその位置に持ってくることが下唇の構造として大切な点となります.
図42.(a) 板状の下唇前基節を持つギンヤンマ.下唇が折り畳まれた状態と,折り畳んだときに大顎が当たる位置.(b) スプーン状の下唇前基節を持つトラフトンボ.下唇が顔の前部を被っている状態と,折り畳んだときに大顎が当たる位置.
ヤンマ科やサナエトンボ科など,板状をしている下唇前基節を持つものの多くは,下唇側片の幅が狭く細長くなっており,可動鉤が非常に長いのが典型的です (図43b).これらにはふつう側刺毛・腮刺毛がありません.下唇を伸ばしこの鋭い可動鉤を使って突き刺すようにして捕獲された餌生物 (図43a)は,下唇をたたむと下唇中片の近くに位置することとなり,大顎で噛み砕かれることになります.
図43.(a) 下唇側片が細長く可動鉤が長く鋭いクロスジギンヤンマが,下唇を伸ばしヌマエビを捕まえた瞬間.(b) カトリヤンマの下唇前基節.下唇側片が細長く可動鉤が長く鋭い.(c) オオキトンボの下唇前基節.スコップのように砂泥をすくい取り,砂とともに餌を捕まえた合成図.実際の採餌行動を観察して,側刺毛・腮刺毛をあとから書き加えて餌を囲い込んだところを示している.側刺毛は下唇側片の付け根に行くほど短くなり,腮刺毛は正中線に近づくほど短くなるのがふつうで,それによって囲いの中央部に穴が開いた状態になり,大顎がそこを通過して餌に近づく.
一方でエゾトンボ科やトンボ科など,深いスプーン状をした下唇前基節を持つものは,下唇側片が三角形状で可動鉤が短く,側刺毛・腮刺毛が長く数が多いのが典型的です.スプーン状の下唇で底の砂泥ごとすくい取られた餌は,側刺毛・腮刺毛を広げて囲い込まれます (図43c).観察していると,下唇をザッザッとザルで砂をゆすぐように動かし,中片や側片の隙間から砂泥を放り出しています.そして砂泥がなくなると大顎で餌を噛み砕きます.こういう動作をする幼虫の左右の下唇側片が接する縁や下唇側片と下唇中片が接する縁には,砂泥は通すが餌は逃がさないように短い毛が生えていることが多いです.
もちろんこれらには種によってさまざまな違いがあります.例えば,イトトンボ科やモノサシトンボ科の下唇前基節は全般的に浅いスプーン状で可動鉤がやや長めです.側刺毛・腮刺毛はありますがトンボ科などに比べると数が少ないものが多いようで,板状とスプーン状の中間的な形態といえます (図44).またオニヤンマ科やミナミヤンマ科の下唇前基節は,特に下唇側片の前縁がするどい歯状になっていて可動鉤も長くなっています (図45).これは下唇側片全体でがっちりと餌生物を挟みこむのでしょう.カワトンボ科の多くは中央欠刻が大きく開いていて,これでどのような種類の餌を捕り口に運ぶのか,想像するのが難しく感じるほどです (図46).
このように下唇の形状は,細かく見ると多様性に富んでいます.なぜこういった形態になっているかについては,下唇の形状と生活場所での主な餌の種類の関係,そしてそれをどう捕獲して食するかといった観察を十分行って考察することがまず出発点になります.しかしこういった体系的観察はほとんど報告がありません.きっとそこからは興味深いトンボたちの生態が浮かび上がってくると思います.
さて,消化器官に入った餌は消化されたあと,糞として肛門から出されます.トンボ幼虫を飼育した方なら経験があると思いますが,幼虫の糞は薄い膜で包まれています.これは中腸の内側の膜で,直腸を通るときに直腸鰓を汚さないようになっています.肛門から排出するときは,かなりの勢いで遠くへ飛ばすことがあります.ヤブヤンマなどでよく観察されますが,これは自身の生活する小さな水たまりを汚さないためであると考えられています(永瀬,1974).小さな容器で飼育していると容器の外へ飛ぶことも珍しくありません.
(1-2) 餌を感知する感覚器官
餌を捕るためには餌を見つけなければなりません.次に餌を感知するいくつかの器官について見ていくことにしましょう.トンボ幼虫は,主に視覚によって餌を見つけていますが,若齢幼虫や砂泥に潜って生活するような幼虫は触覚や振動を感じる機械受容器を主に用いているようです.また化学受容器もあることが示唆されています (Corbet, 1999).
まず視覚から見ていきましょう.前幼虫から脱皮した2齢幼虫の複眼には,例えばアオナガイトトンボ新村 (2002) では個眼が7つと記載されています.他の前幼虫・2例幼虫の写真を見ても多くは個眼は7つです.これは脱皮成長する度に数が増えていきます.ヤンマ科の
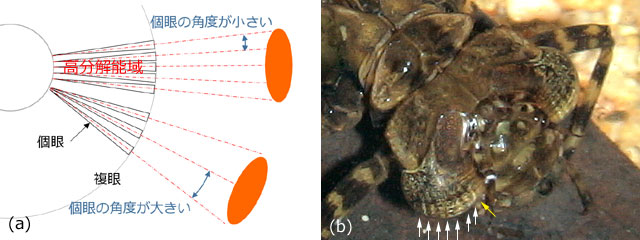
Aeshna palmata を使って調べられた研究では (Sherk, 1977),個眼は前方で新たにつくられ,古い個眼は後方へ移動していきます.前方で形成された新しい個眼の視線角度は非常に小さく,そのあたりの個眼はまとまってほぼ一つの方向を見ていることになります.その結果その領域の分解能が高くなります (図47(a)).これを高分解能域と呼んでいます.
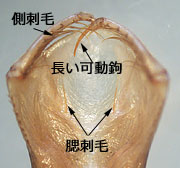
図44.キイトトンボの下唇前基節 (一部).側刺毛と腮刺毛があるが可動鉤は鋭く長い.全体は下唇側片がねじれの位置にあり浅いスプーン状を呈する.腮刺毛が1本しかなく,餌の囲い込みはできないだろう.どのように餌を捕獲するのか,想像が難しい.
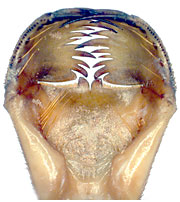
図45.ミナミヤンマの下唇前基節 (一部).典型的なスプーン状の下唇前基節だが,下唇側片の前縁は鋭い歯状になっており,可動鉤も長い.
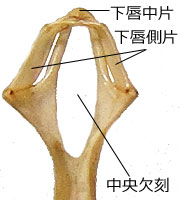
図45.ミヤマカワトンボの下唇前基節 (一部).浅いスプーン状の下唇前基節といえるが,中央欠刻が異常に大きく下唇中片先端部が下唇側片より前に出ている.下唇側片を少し開くとちょうど下唇中片先端部ぐらいの位置になるようだ.
図47.(a) 個眼の角度と高分解能域の説明図.個眼の視線角度が小さければ,図では物体を5つの個眼で見ていることになるが,個眼の視線角度が大きければ,同じ物体を3つの個眼で見ることになり,分解能(解像度)が落ちる.(b) 黒いスジになって見える擬瞳孔は,視線角度が小さい個眼の集まった部分である.黄色矢印のあたりで脱皮時に形成されたこの個眼の集まりは,脱皮ごとに後ろへ順次移動していくので(白色矢印),このようなスジが見える.これは一種の年輪といえる.(ミルンヤンマ終齢幼虫).Sherk (1977) を参照した.
個眼の視線角度が小さくなると,そこへ入った光が吸収され反射してこなくなるので,複眼を見ている我々には黒く見えてしまいます.これが擬瞳孔です.擬瞳孔が大きければ,それだけ多くの個眼が同じような方向を向いている証拠となり,トンボにとっては,その方向が高解像度でよく見えているということになります (欄外図48).図47(b)のミルンヤンマではスジ状の擬瞳孔が見られますが,このようなスジはすべてのトンボに見られるというわけではありません.
図49.餌の方に向かって定位し高分解能域で餌との距離を測るヤンマの幼虫 (模式図).
トンボの眼球はヒトのように動かないのでまず体 (頭部) を餌の方に向ける.前方の高分解能域に存在する複数の個眼は,それぞれの視線角度が小さくほぼ同じ方向を見ていることになる (図47).そして両眼のそれら領域の個眼視線が交わる位置が下唇のとどく位置に一致している.
次に機械受容器を見てみましょう.Rowe (1994) は,イトトンボ科の
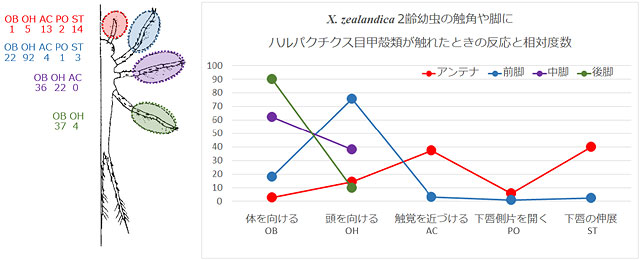
Xanthocnemis zealandica の2齢幼虫を使って,機械受容器の実験をしています.アンテナ (触角),前・中・後脚の各脛節より先の部分に,ハルパクチクス目甲殻類が当たったときの幼虫の反応を見ています (図50).2齢幼虫はまだ複眼の発達が悪く,主に機械受容器で餌を捕っていると考えられています.またハルパクチクスとはケンミジンコとよく似た形態をした甲殻類です.
図50のグラフから,中脚・後脚に触れると体を向ける,前脚に触れると頭を向ける,触角に触れると下唇を伸ばす,といった反応がもっとも多く出ています.このことから,これらの部位は,餌生物を認知する機械的受容器の役割を果たしていることが分かります.トンボの目は動きませんので体を餌の方に向ける必要がありますから,餌が脚に触れたときは必ず体や頭を向ける反応が出ています.そして触角に当たったときは下唇の動きが出て捕獲動作に移行するのが分かります.
図50.ハルパクチルス目甲殻類が触れたときの反応 (模式図).Rowe (1994) より.
脛節より先の脚には毛がある.これは電子顕微鏡写真より機械受容器と想定している.Rowe (1994) では幼虫の行動に13のカテゴリーを認めており,そのうちの5つの行動(OB, OH, AC, PO, ST)が見られたという.グラフはオリジナル.幼虫の絵は Rowe (1994) に色をつけた.
若齢幼虫には体表にさまざまな突起物が見られることがあります (Corbet, 1962).また渡辺 (1995;1996)はメガネサナエ属の2齢幼虫の腹部背面に2列の突起物があることを報告しています.これらは機械受容器なのかもしれません.また砂泥中に潜って餌を待つタイプの幼虫は,機械受容器が重要な役割を果たしているようです.

図48.成虫の擬瞳孔.
緑色の複眼の中に見える黒い円が擬瞳孔.視線が同じ方向を向いている個眼が集まっている部分.高分解能域でもありその方向の物体は緻密に見える.前方の擬瞳孔が大きいことから,前方がよく見えていることが分かる (ヒメサナエの成虫).
トンボの生態学 4.幼虫の生活
幼虫の微小生息場所
さて,温度,呼吸,捕食者,採餌と,トンボ幼虫の生存にとって重要な要素についてお話をしてきました.温度はトンボの分布を決めるという点で他とは少し異なりますが,それ以外の要素は,系統的な制約のもとで,トンボの生息場所や微小生息場所を決める選択圧となるでしょう.言いかえると,トンボ自身が生まれつき持っている能力 (系統的制約) を使って,捕食をうまく回避でき (捕食回避),採餌が最適に行える (最適採餌) ような場所は,その幼虫が生き残りやすい場所ですから (繁殖成功度に寄与),現在の幼虫の微小生息場所は,進化の産物であるという考え方になります.
さて,幼虫の微小生息場所選択,これは幼虫の生活形態と言いかえることもできます.これについては,今までにいくつかの分類が試みられています.例えば枝 (1976) は,それまでの研究者の用語を整理して3つに分類しています.遊離型,地面型,地下型です.遊離型とは水底から離れて生活するという意味で,水生植物や朽ち木・岩などにつかまって生活しているタイプです.地面型とは水底で生活するが潜らないという意味で,地下型とは水底で生活して沈積物や砂泥に潜って (隠れて) 生活するという意味になります.また杉村ら (1999) は,まず水中で生活するタイプ (遊離型に相当) と水底で生活するタイプに分け,後者をさらに這い回り型 (地面型に相当) と浅く潜るタイプ (地下型に相当) に分けて,基本的には同様の生活形態分類をしています.
ただしこれら研究者も指摘していますが,幼虫の生活形態は,状況によって二つ以上にまたがることがあり,種ごとにどれかのカテゴリーに分類することには無理があります.例えば夏の間沈水植物内で暮らしているクロイトトンボ幼虫は (遊離型),冬に水生植物が枯れてなくなると,水底沈積物の間に紛れ込んで生活します (地面型).
Corbet (1999) は,上記のように幼虫の生活形態をカテゴリー分けすることの困難さを意識しつつも,トンボ幼虫の行動・形態の多様性 (系統的制約の種ごとの多様性) が微小生息場所と関係している点に注目して,次のようなカテゴリー分けを提案しています.不均翅亜目・ムカシトンボ亜目については,静止時の脚の使い方をもとに,(1) しがみつきタイプ,(2) 腹ばいタイプ,(3) 潜伏タイプ,(4) 穴掘りタイプ,の4つに分けています.また均翅亜目については,尾鰓の形態をもとに,(1) 胞状または三稜状,(2) 三稜状ないし葉状,(3) 水平な薄葉状,(4) 垂直な薄葉状,の4つです.これは尾鰓の形状が微小生息場所と比較的よく対応しているからです.
ただいずれの分類も,底生生活をする幼虫に関して,深く潜っている,浅く潜っている,沈積物に埋もれているだけ,潜っていなくて泥が背面に付着しているだけ,等の区別が,野外での生活状態が継続的に観察できないので,実際のところ判別が難しく,いつも迷ってしまいます.そこで,潜るかどうかを分類基準から外し,底生型とまとめてしまうことで,かなりすっきりとした分類ができると考えています.
そして,幼虫がもっとも生存確率の高くなるような場所に成虫が産卵するように進化が起きているはずだと考えて,成虫の産卵様式と産卵基質を大きな枠組みとして用いると,原則的にうまくまとまるように思えます (表1).もちろん表1は,その分類群の大多数がそのカテゴリーに入るということで,例外的な微小生息場所を選ぶ種はいくつか存在します.また季節的に,また状況に応じて,微小生息場所を変える種もふつうにいますので,あくまで成長が盛んな活動期における,その分類群の典型的な微小生息場所と考えてください.
|
| 産卵様式 |
産卵基質 |
生活場所 |
微小生息場所 |
分類群 |
| 植物内産卵 |
水生植物 |
水生植物依存的 |
遊離型 |
アオイトトンボ科 |
| (産卵管) |
(抽水) |
|
|
カワトンボ科(一部) |
|
(浮葉) |
|
|
イトトンボ科 |
|
(沈水) |
|
|
ヤンマ科(一部) |
|
|
|
底生型 |
モノサシ属・グンバイ属 |
|
水生植物外 |
水生植物非依存的 |
遊離型 |
ミナミカワトンボ科 |
|
(陸上植物) |
|
|
ムカシトンボ科 |
|
(朽木) |
|
底生型 |
ルリモントンボ属 |
|
(土中) |
|
|
ヤンマ科(一部) |
|
|
|
|
カワトンボ科(一部) |
|
|
|
|
ハナダカトンボ科 |
|
(苔湿土) |
|
その他自由水 |
トゲオトンボ科 |
|
|
|
|
ムカシヤンマ科 |
| 植物外産卵 |
(水面) |
|
底生型 |
サナエトンボ科 |
| (産卵弁) |
(泥面) |
|
|
ミナミヤンマ科 |
|
(浮遊物) |
|
|
オニヤンマ科 |
|
(陸上) |
|
|
エゾトンボ科 |
|
(植物表面) |
|
|
ヤマトンボ科 |
|
|
|
|
トンボ科(一部除く) |
|
|
|
その他自由水 |
キイロハラビロトンボ |
表1.日本産のトンボに関して,産卵様式と産卵基質を大きなカテゴリーとしてまとめると,原則的に,水生植物に依存した生活場所を選ぶ幼虫と,水生植物に特に依存していない生活場所を選ぶ幼虫に分かれる.あとはそれぞれについて,遊離型と底生型に分けると分類がうまくいくように思われる.ただし,浸出水のある場所に生息しているトゲオトンボ科やムカシヤンマ科は遊離型,底生型の分類になじまないこと,また熱帯性のトンボにいくつか存在するファイトテルマータで生活するトンボは,国内にキイロハラビロトンボのみが知られているので,これらはその他の自由水として別扱いとした.
|
それでは,各微小生息場所のカテゴリーについて,代表的な種とともに解説していきましょう.ただ兵庫県に分布しないトンボについては,わずかな観察例や図鑑などの記述をもとにしています.
カテゴリー1.植物内産卵-水生植物依存的遊離型
図51.ヒヌマイトトンボ幼虫とギンヤンマ幼虫(飼育).
ヒヌマイトトンボはヨシの植生内で,枯れたヨシの茎などにつかまって生活している.ギンヤンマは緑色の植生内で脱皮すると,体色がこのように緑色の隠蔽色になる.
このカテゴリーのトンボの卵は生きた水生植物内に産み付けられることがふつうで,孵化した幼虫は原則として産み付けられた植生内で生活していると言ってよいでしょう.体は細長く,水生植物の茎にしがみつけば遠目には一体化し,隠蔽されます (図51左).ギンヤンマの体色などは緑色のものも多いです (図51右).水面近くの水生植生内は明るいので,視覚で餌を捕ります.そのため複眼が大きく発達しています (図51右の複眼).
下唇前基節は長いものが多く (欄外図52),動いて餌を探し,少し離れた位置からでも餌を捕らえることができます.ヤンマ科は板状の下唇前基節で,下唇側片の可動鉤ではさんで餌を捕ります (図43a).アオイトトンボ科では,下唇前基節は浅いスプーン状ですが,可動鉤が長く下唇側片の前縁が鋭い歯状になっています (欄外図53).おそらく下唇側片全体ではさむようにして餌を捕るのでしょう.これはヤンマ科に近い餌の捕り方で,一種の収斂と考えられます.植生内はコミズムシ (フウセンムシ) のような小さな昆虫が泳いできますから,こういった小昆虫を捕るのに適しているように思えます.
それに対してイトトンボ科の下唇はそれほど長くなく,側刺毛や腮刺毛が発達しているものが多いです.また下唇側片もアオイトトンボほど刺々しくありません.これはトンボ科などと同じく,すくい取り法で餌を捕るのでしょう.植生内は,ミジンコ,ケンミジンコ,カイミジンコ類が結構たくさんいますので,こういった微小な甲殻類を捕獲するのに適したつくりかと思われます.これらは,同じ植生内生活者として,餌を異にすることで,種間競争を低減させているのかも知れません.なおアオイトトンボ科の下唇前基節にも側刺毛や腮刺毛があり,ヤンマ的な狩猟の他にイトトンボ科的な狩猟もできる,両用の下唇前基節に見えます.
日本産のイトトンボ科とアオイトトンボ科は,すべてこのカテゴリーに入れてよいように思えます.カワトンボ科ではアオハダトンボ,ハグロトンボがこのカテゴリーに入りますが,根際で泥にまみれて生活している姿を見かけます (図34左).ヤンマ科では,ギンヤンマ属,マルタンヤンマ,マダラヤンマ,オオルリボシヤンマなどはここに含まれるでしょう.このカテゴリーの幼虫は,水生植物が枯れると底生生活者になり,落ち葉の間などで過ごすのがふつうです.
カテゴリー2.植物内産卵-水生植物依存的底生型
図54.モノサシトンボ幼虫とグンバイトンボ幼虫(飼育).
モノサシトンボ幼虫は,休息時に脚が長く横に広がって扁平な姿勢である.これは落ち葉などの隙間で生活する潜伏タイプ幼虫の特徴である.グンバイトンボ幼虫も似ているが,モノサシトンボほど横に広がらないので,これはカテゴリー1.に含めるべきかも知れない.
このカテゴリーのトンボの卵も生きた水生植物内に産み付けられます.カテゴリー1.と異なる点は,潜伏タイプの特徴である,長い脚を横に広げて休止する姿勢をとります.これは植生内というよりむしろ底生生活者の特徴です.また大きな尾鰓は,溶存酸素が少なくなりやすい水底への適応と考えられます.そういう理由でこのカテゴリーを設けました.
カテゴリー3.植物内産卵-水生植物非依存的遊離型
図55.コナカハグロトンボ幼虫 (標本) とムカシトンボ幼虫 (野外).
コナカハグロトンボ幼虫は尾鰓が胞状で吸着性がある.流れの石の表面にはりついてその上を這い回って生活している.ムカシトンボ幼虫が石の表面に吸着している.
このカテゴリーのトンボの産卵方式は植物内産卵ですが,ふつう生きた水生植物ではなく朽ち木や陸上植物に産卵するトンボです.必ずしも水生植物があるところに産卵するわけではないので,幼虫は水生植物非依存的な生活を送ります.その中でもこのカテゴリーの幼虫は水底を利用することなく,瀬石などにはりついて底より上の部分で生活している幼虫たちです.もちろん底との隙間に入り込むと水底の位置に潜むことになりますが,腹面が底ではなく石の方に向いている点で底質を利用していないことになります.いずれも流水性種です.また接触走性が強いトンボたちです.
ミナミカワトンボ科にはコナカハグロトンボ以外にチビカワトンボがいます.産卵は朽ち木や苔に行われているようで,やはり胞状の尾鰓があります.ムカシトンボは流畔の陸生植物に産卵します.下唇前基節はいずれも板状です.堅い基質にはりついた状態で下唇を伸ばして餌を捕ろうとすると,板状の方がうまくいくように思えます.
カテゴリー4.植物内産卵-水生植物非依存的底生型
図56.ミヤマカワトンボ幼虫とミルンヤンマ幼虫 (飼育).
いずれも水生植物のあまり生えない河川の上流部に棲息しており,淵などの底に堆積した落ち葉などの中に潜っている.
このカテゴリーのトンボの産卵方式は植物内産卵です.ふつう朽ち木に産卵します.産卵場所に水生植物がないことが多い,細流や上流に棲息するトンボが中心です.幼虫は水底で生活していますが,水底の基質に潜ることは基本的にはありません.落ち葉などの間に潜伏するタイプです.
南西諸島に見られるルリモントンボ属は,源流近くの細流に多く見られ,朽ち木に産卵し,幼虫は浅い流れで生活しています.カワトンボ科ではアサヒナカワトンボ,クロイワカワトンボ,ミヤマカワトンボ,リュウキュウハグロトンボが,主に朽ち木や湿土に産卵し,上流域の石がゴロゴロしたような流れで生活しています.ニホンカワトンボはやや中流域の抽水植物の生えているところによく見られますが,主に朽ち木を選んで産卵しています.これらの脚は横に平たく広がり,潜伏タイプの特徴です.ハナダカトンボ科も同様です.ヤンマ科では朽ち木や土中に産卵する,ミルンヤンマ,コシボソヤンマ,ヤブヤンマなどが含まれます.サラサヤンマは朽ち木や湿土に産卵し,湿地の水たまりやときには湿った落ち葉の間などで生活していますが,このカテゴリーに入れてもよいでしょう.
もしこれらの生息場所に水生植物が繁茂しているとそれを利用して隠れていることもありますが,水生植物がない場所でふつうに生活していますので,基本的に水生植物非依存的底生型の幼虫と考えるべきでしょう.
カテゴリー5.植物内産卵-水生植物非依存的その他自由水型
図57.ヤンバルトゲオトンボ幼虫とムカシヤンマ幼虫 (野外).
浸出水のあるところに生育するトンボたちで,産卵はそこに生えている陸上植物,苔,湿土などに産卵する.
このカテゴリーのトンボは,やや特殊な環境に生育するトンボたちです.ふつうの河川や池沼ではなく,浸出水のあるところで生活しています.産卵は朽ち木,苔,湿土に行われます.トゲオトンボ科は岩の表面 (図57左) や落ち葉の間に潜んでおり,ムカシヤンマは穴を掘ってその中で暮らしています (図57右).トゲオトンボ科は河川の上流域で,川に流れ込む浸出水に見られることがあるので,上流域のトンボに見えるかも知れませんが,ふつう幼虫は本流の流れでは生活していません.
海外では,ファイトテルマータ (樹洞その他) に棲息する,ハビロイトトンボ科などが見られますが,日本国内では,産卵管を持つトンボでファイトテルマータに棲息するトンボは知られていません.
カテゴリー6.植物外産卵-水生植物非依存的底生型
図58.タベサナエ幼虫とタイワンコヤマトンボ幼虫 (野外).
図59.オニヤンマ幼虫 (野外) とウスバキトンボ幼虫 (飼育).
植物外産卵,いわゆる打水産卵,打泥産卵,空中産卵などを行う種群である.産卵弁を持って産卵するトンボのほとんどはこのカテゴリーに入る.
このカテゴリーは数多くの種群を含みます.産卵弁を持っていて植物内産卵を行わない種のほとんどはこのカテゴリーに入ります.均翅亜目はすべて産卵管を持ちますから,すべて不均翅亜目になります.
トンボ目の系統について考察したページを参照していただくと分かりますが,どの研究者の不均翅亜目の系統樹を見ても,産卵管を持つヤンマ科が古くに分岐し,産卵弁を持つ系統はその後に出現しています.
産卵弁を持つことで,産卵時に水生植物や朽ち木などに依存する必要がなくなり,さまざまな水域へ適応放散していくことができました.一方その結果,水生植物がないところでは,遊離型という水底から離れた生活場所が原理的になくなりますので,必然的に底生生活になってしまいます.
この中には,Corbet (1999) の分類にある,腹ばいタイプ,潜伏タイプ,穴掘りタイプのすべてが含まれます.上で紹介した図58,59の写真を例にとると,トンボ科のウスバキトンボは腹ばいタイプで,ヤマトンボ科のタイワンコヤマトンボは潜伏タイプ,そしてサナエトンボ科のタベサナエ,オニヤンマ科のオニヤンマなどは穴掘りタイプに分類されます.
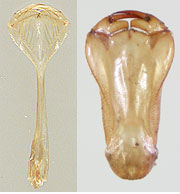
図52.カテゴリー1.の下唇前基節.長いものが多い.左はアオイトトンボ,右はオオルリボシヤンマ.アオイトトンボの下唇は浅いスプーン状で下唇側片の先端部が鋭い歯状になっている.

図53.アオイトトンボの下唇側片.先が鋭く切れて歯状になっている.
下唇前基節の形状で見ると,サナエトンボ科だけが板状で,他はスプーン状です.サナエトンボ科の大部分は穴掘りタイプですが,潜った状態で下唇を伸展させるためには,長さの短い薄い板状の方が底の砂泥などの抵抗を受けにくいと思われますので,適応的形質だと思われます (欄外図60左).しかし同じ穴掘りタイプと思われるオニヤンマ科やミナミヤンマ科は大きく深いスプーン状です (欄外図60右).しかしこれらの下唇側片の前縁部には深い切れ込みがあります.カテゴリー1.のアオイトトンボ科で指摘したときと同様の状況が見て取れます.つまり下唇側片全体で挟みこむ捕獲方法です.
オニヤンマ科やミナミヤンマ科は,図59左の写真の体を反らしている姿勢のように,底に浅く潜り,頭と腹部先端を水底から出し,呼吸と採餌を行っているようです.そしてその上を通った餌にがぶりと噛みつくのでしょう.これには実際に野外での観察経験があります.川にできた水たまりで幼虫採集をしましたら,いくつかの大型の幼虫が採れました.オニヤンマは同定が楽ですので先に放したら,それはすぐに砂底に潜りました.そのあと同定困難なオオルリボシヤンマとルリボシヤンマの種名確定を慎重に行った後,オオルリボシヤンマの終齢幼虫を放しました.するとそれが少し水底を歩いたときに,先のオニヤンマにがぶりと捕らえられてしまいました.
オニヤンマ科やミナミヤンマ科の下唇前基節には側刺毛や腮刺毛があり,トンボ科などのスプーン状の下唇前基節と同様の,底の砂をすくって餌捕獲をする機能もあると思われます.
このカテゴリー6.には多くの種群が含まれていますが,腹ばいタイプ,潜伏タイプ,穴掘りタイプと,微小生息場所を分かつことでニッチを分け合い,水底という生息場所で種間競争を軽減しつつ共存を果たしているのでしょう.
カテゴリー7.植物外産卵-水生植物非依存的その他自由水型
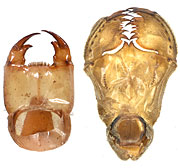
図60.ヤマサナエの下唇前基節(左)と,オニヤンマの下唇前基節(右).
日本国内で唯一知られているファイトテルマータに棲息するトンボがいます.キイロハラビロトンボです.台湾のトンボ図説 (汪,2000) には,「幼虫の生息場所は特殊で,樹洞,竹の穴,黒っぽいコンテナなどである.」との記述があり,このトンボの台湾名が,なんと「樹穴蜻蛉*14」という名だそうです.
以上,成虫の産卵様式と産卵基質を軸にまとめてみました.みなさんも,幼虫の微小生息場所について,いろいろと考えていただけると幸いです.以上で,幼虫の生活に関する章を終わります.
*14.「蜻蛉」ではなく「蜻+虫へんに廷のつくり」の文字で書かれている.Unicodeになるので「蛉」の字を当てた
トンボの生態学 5.幼虫の生活
参考文献
*Åbro, A., 1974. The gregarine infection in different species of Odonata from the same habitat. Zool. Scripta 3:111-120.
浅島 誠,ほか27名,2020.改訂生物.2・東書生物306.東京書籍.
新井 裕,1987.トンボ幼虫の擬死.インセクタリウム, 24(12):358-361.
東 輝弥・青木典司,2025.兵庫のトンボ.自刊.
Corbet, P.S., 1962. A Biology of Dragonflies. H. F. & G. Witherby ltd., GB.
Corbet, P. S., 1999. Dragonflies Behavior and Ecology of Odonata. Cornell Universitiy Press. New York.
枝 重夫,1976.トンボの採集と観察.ニュー・サイエンス社 グリーンブックス22.東京.
後閑暢夫,1984.昆虫の体制.昆虫の生物学[第二版]:47-60. 玉川大学出版部,東京.
広瀬良宏・伊藤智・横山透,2007.北海道のトンボ図鑑.いかだ社,東京.
Johansson, F., & Samuelsson, L., 1994. Fish-induced variation in abdominal spine length of Leucorrhinia dubia (Odonata) larvae? Oecologia 100:74-49. (English Abstract 9993 in Odonatologica).
*Jones, S.P., 1985.A note on the survival of dragonflies in adverse conditions in Cornwall. JBDS 1:83-84. (in Corbet, 1999).
Komnick, H., 1978. Osmoregulatory role and transport ATPase of the rectum of dragonfly larvae. Odonatologica 7:247-262.
*Miller, P. L., 1993. Responses of rectal pumping to oxygen lack by larval Calopteryx splendens (Zygoptera: Odonata). Physiol. Entomol. NtO 4:6-8.
Miyatake, T, K. Katayamay, Y. Takeda, A. Nakashima, A. Sugita & M. Mizumoto, 2004. Is death-feigning adaptive? Heritable variation in fitness difference of death-feigning behaviour. Proc. R. Soc. Lond. B, 271:2293-2296.
永瀬幸一,1974.ヤブヤンマにおける Project pellets と Cannibalism.Gracile (16):6-7.
Pavlyuk, R. S., 1978. Structure of dragonfly parasitocoenoses (Odonata, Insecta) from various aquatic habitats of the western Ukraine. IR. Tezisy Doki. I. Vses. S'ezd. Parazitocenol, 2:38-40. (English Abstract 2385 in Odonatologica).
Rowe,R.J., 1994.Predatory behaviour and predatory versatility in young larvae of the dragonfly Xanthocnemis zealandica (Odonata: Coenagrionidae).
New Zealand Journal of Zoology 21:151-166. (https://www.tandfonline.com/doi/pdf/10.1080/03014223.1994.9517983)
*Sato, T., Watanabe, K., Kanaiwa, M., Niizuma, Y., Harada, Y. and Lafferty, K.D. 2011. Nematomorph parasites drive energy flow through a riparian ecosystem. Ecology 92:201-207.
*Schott, R. J., & & Brusven, M. A., 1980. The ecology and electrophoretic analysis of the damselfly, Argia vivida Hagen, living in a geothermal gradient. Hydrobiologia 69:261-265.
*Sherk, T. E., 1977. Development of the compound eyes of dragonflies (Odonata) I. Larval compound eyes. J. Exp. Zool. 201:392-416.
新村捷介,2002.アオナガイトトンボの卵.Gracile (64):1-2.
杉村光俊・石田昇三・小島圭三・石田勝義・青木典司,1999.原色日本トンボ幼虫成虫大図鑑.北海道大学図書刊行会.札幌.
*Thorp, J. H., & Diggins, M. R., 1982, Factors affecting depth distribution of dragonflies and other benthic insects in a thermally destabilized reservoir. Hydrobiologia 87:33-44.
Tillyard, R. J., 1917. The biology of Odonata (Odonata or Paraneuroptera). Cambridge University Press. London.
汪 良沖,2000.台灣的蜻蛉.349pp.,人人月暦股イ分有限公司,台北.
渡辺庸子,1995.メガネサナエとナゴヤサナエの卵と1,2齢幼虫の形態.Tombo 38(1/4):56-57.
渡辺庸子,1996.オオサカサナエの卵と1齢幼虫について.Tombo 39(1/4):44-45.