�g���{�̐��Ԋw
���m�O���t�F�A�J�g���{�@�ڎ�
�g���{�̐��Ԋw�@���m�O���t�F�A�J�g���{
�A�J�g���{�ɂ���
�@�u�A�J�g���{�v�́C�����ʂ�ł��Ɓu�ԂƂ�ځv�C�܂�Ԃ��g���{�Ƃ������ƂɂȂ�܂��D�Ԃ��̐F�̃g���{�́C�g�߂ɂ����\�������̎�ނ����܂��D�V���E�W���E�g���{�C�n�b�`���E�g���{�C�n�l�r���g���{�C�x�j�g���{���͂��߁C������֍s���ƁC�I�I�n���r���g���{�C�E�~�A�J�g���{�Ƃ������̂܂ł��܂��D�����́C���ɃI�X�ɂ����āC���炾���Ԃ��F�����Ă��܂��D�u�A�J�g���{�v�Ƃ����ꎩ�̂����ފw�Ɏg���Ă�����{�ꖼ�̂ł͂Ȃ��C�����鑭�̂ł�����C�u�A�J�g���{�v�Ƃ������̂��ǂ̂悤�ȃg���{�Ɏg�����ƈ���ɂ��܂�Ȃ��Ƃ������ƂɂȂ�܂��D
�@�ł́C���ފw�Ŏg���Ă���a���Łu�A�J�g���{�v���Ӑ}������{��͉����Ƃ����܂��ƁC����́u�A�J�l��*1�v�ł��D�w���ł� Genus Sympetrum Newman, 1833 �Ə�����܂��D�P�� Sympetrum (�V���y�g����)�ł����܂��܂���D�����Łu�Ӑ}����v�Ə������̂́C�������̑������u�ԂƂ�ځv�ŃC���[�W����̂́C�O�ؘI���̓��w�u�ԂƂ�ځv�̉̎��ɏo�Ă���u�ԂƂ�ځv�ł͂Ȃ����Ǝv���邩��ł��D
�@���́u�ԂƂ�ځv�̐��̂ɂ��Ă͏��������āC�A�J�l���̃A�L�A�J�l���낤�Ƃ����̂���ʓI(�Ⴆ�C����C1988)�ł����C�A�J�l���ł͂Ȃ��E�X�o�L�g���{�ł���ƍl���Ă���l������悤�ł�(�Ⴆ�ΐ����E���C1990)�D����(1988)�́C�u�ԂƂ�ڎ~�܂��Ă����Ƃ̐�v�Ƃ������w�u�ԂƂ�ځv�̌��_�ƂȂ��Ă���O�ؘI���̔o������������ɏo���āC�A�L�A�J�l���L�͂ł���Əq�ׂĂ��܂��D�m���ɃE�X�o�L�g���{�͊Ƃ̐�ɂ͎~�܂�܂���D����C�����E���(1990)�́C�u�[�Ă����Ă̐ԂƂ�ځv�́u�[�Ă��v�̎����ɂ̓A�L�A�J�l�͌Q���E�X�o�L�g���{�Ȃ�Q��Ƃ������Ƃ������ɂ��Ă��܂��D
�@�����b������Ă��܂����悤�ȋC�����܂��D�u�ԂƂ�ځv�ɂ��Ă̋c�_�͂��Ă����C���̃y�[�W�ł͕��ފw��̃A�J�l�����������ƍl���Ă��܂��̂ŁC�ȍ~�A�J�g���{�Ƃ�������g�킸�C�u�A�J�l���v�Ƃ�����Řb��i�߂Ă��������Ǝv���܂��D
*1. ���� Sympetrum �̘a���ɂ��āC�l�c�E���(1985)�ł́u�A�J�g���{���v��p���Ă���D�܂�������(1999)�ł́u�A�J�g���{�i�A�J�l�j���v�C�Γc��(1988)�ł́C�u�A�J�l�i�A�J�g���{�j���v�ƋL�q���Ă���D���̑��̂��낢��ȏo�ŕ��ׂĂ݂�ƁC�A�J�l���ƃA�J�g���{���̗�����������D�����ł́C������(2012)�ɂ��������āu�A�J�l���v��p���Ă���D
�ʐ^�P�D�Ԃ�����ǂ����ފw�I�Ɂu�A�J�l���v�ł͂Ȃ��g���{�����D
�@��i������F�V���E�W���E�g���{�C�n�b�`���E�g���{�C�n�l�r���g���{�C
���i������F�x�j�g���{�C�I�I�n���r���g���{�C�E�~�A�J�g���{�D
�@�ʐ^�P��6��̃g���{�͂��炾���Ԃ��F�����Ă��܂����C���ׂăA�J�l���ł͂���܂���D�A�J�l���̃g���{�͓��{�̃g���{�Ȃ̒��ł͈�ԑ傫�ȑ����`�����Ă��āC���Ɍ��ɂ�15��(���{�����ł�17��1����)���������Ă��܂�*2�D�A�J�l���̒��ɂ͑嗤��������������āC2018�N1�����݃A�J�l���ł́C�^�C���N�A�L�A�J�l�C�I�i�K�A�J�l�C�X�i�A�J�l��3�킪���Ɍ��ŋL�^����Ă��܂��D
�@�A�J�l���̃g���{�͈�ʂɑ̐F���Ԃ��Ȃ���̂������̂ł����C�ꕔ�I�X�E���X�Ƃ��̐F���Ԃ��Ȃ�Ȃ����̂�����܂��D���{�Y�̂��̂ł́C�i�j���g���{�C�}�_���i�j���g���{�C���c�A�J�l�������ł��D�����́C���A�J�g���{�Ƃ������A�J�g���{�Ƃ������邱�Ƃ�����܂��D
*2. ��܂Ŋ܂߂�ƁC�����ɋL�^�̂���A�J�l���̃g���{��21��1����ƂȂ�D���̒�����C�X�i�A�J�l�C�^�C���N�A�L�A�J�l�C�}���V���E�A�J�l�C�I�i�K�A�J�l�������āC17��1���킪�������Ă���D����ɕ��Ɍ��ł́C�G�]�A�J�l�C���c�A�J�l�C�q�����X�A�J�l(���X�A�J�l�̖k�C������)��������āC15��ƂȂ�D
�ʐ^�Q�D�Ԃ��Ȃ�Ȃ��A�J�l���̃g���{�����D������F�i�j���g���{�C�}�_���i�j���g���{�C���c�A�J�l�D��������I�X�D
�@�ł́C�A�J�l���̃g���{�Ƃ����̂́C�ǂ����������������g���{�̏W�܂�Ȃ̂ł��傤���D�A�J�l���̃g���{���Ђƌ��Œ�`����͎̂��͔��ɓ���̂ł��D����ɂ��ẮC�������I�ɂȂ�܂����C�������g�����������ʂ��Љ�܂��傤�D
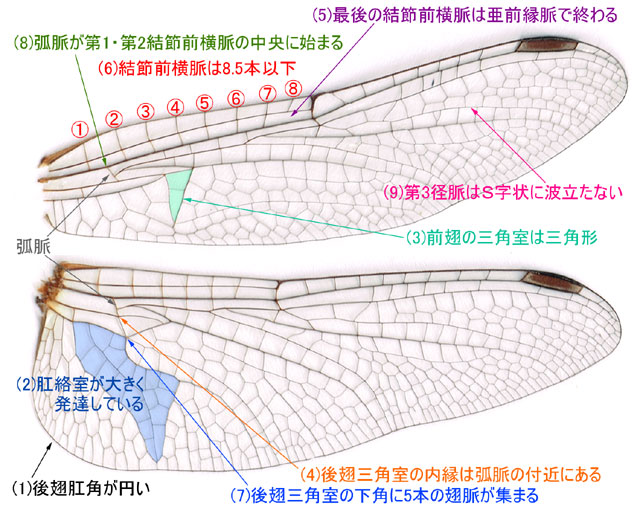
�}�P�D�A�J�l���������̓��� (Fraser�C1957�G���C2010�G�l�c�E���C1985)�D
�i�c�A�J�l�� �D
�@�}�P���g���Č�������ꍇ�́C�ʏ�C(1)����(9)�̏��ɂ݂Ă����Ƃ悢�ł��傤�D������(8)(9)�Ȃǂ͔��ɔ����ŁC�q���L�g���{�Ƃ�������ɂ���g���{�ł́u(8)���������͂���1���ߑO�������ŁC(9)���A�J�l�����͂��g�ł��Ă��銴���v�Ƃ����킸���ȈႢ��������܂���(���C1985)�D���ꂾ�������ɓ�������ׂĂ�(����I��`)�C�A�J�l�����`����͓̂���Ƃ����̂����ۂ̂Ƃ���ł��D�����Ŏ��ɁC���Ɍ��암�ɍi���āC�A�J�l���̃g���{�����ׂďЉ�邱�Ƃɂ��܂��傤(�O���I��`)�D
�g���{�̐��Ԋw�@���m�O���t�F�A�J�g���{
���Ɍ��Ō�����A�J�l���̃g���{����
�@���Ɍ��ɂ́C���łɏq�ׂ��悤�ɁC15��̃A�J�l���̃g���{���������Ă��܂��D���̂Ƃ����ł�����͂���܂��C��Ŋ뜜�T�ނ̃}�_���i�j���g���{�����O�̓��ɂȂ��Ă��܂��D�S���I�Ɍ������Ă���C��������Ŋ뜜�T�ނ̃I�I�L�g���{�C��Ŋ뜜�U�ނ̃i�j���g���{�̕��́C���Ɍ��암�n���ɂ͂܂������������c���Ă��܂��D�ł́C�������₷�������珇�ɏЉ�Ă������Ƃɂ��܂��傤�D���L�ȊO�̔�ɂ��ẮC�u���Ɍ��Ƃ��̋ߗׂ̃g���{�����v���������������D
�ʐ^�R�D���ɂ�������I�ȃo���h�ł����ɕ�����D�c���͏����p���H�Ȃǂ̗����Ő������Ă���D
�ʐ^�S�D���̊�ɃI�����W�F���L����g���{�ł���H���r�߂��̍����̂Ă���Ɏ~�܂��Ă��邱�Ƃ������D
�ʐ^�T�D���̊����O���ɂ����ăI�����W�F���L���L����D�H�x���Ɍ��ꎞ�ɂ͔N���z���Ă������邱�Ƃ�����D
�ʐ^�U�D��Ŋ뜜�T�ށD9�����{���琅���������r�Ɍ����D10�����낪�ł������D�V�n���Ă���ƐԒ��F�ɂȂ�D
�ʐ^�V�D���̐�[�Ɋ��F��(�m�V����)������3��̂����̈�D�I�X�͕����݂̂��Ԃ��Ȃ�D
�ʐ^�W�D�m�V����������A�J�l���̃g���{�̂����ő�D�w���̔���͔�є�тɑ����C���X�A�J�l�̂悤�ɂׂ����Ƃ��Ă��Ȃ��D
�ʐ^�X�D�m�V�����͔Z���͂�����Ƃ��Ă���D�I�X�͋������ʂ܂ŐԂ��Ȃ�D�v�[���Ȃǂɂ��悭�����D
�ʐ^�P�O�D�I�X�͐Ԃ��Ȃ炸�C�������𐁂����悤�ɂȂ�D7����{�ɂ͒r�̖؉A�Ɏp�������Ă���D
�ʐ^�P�P�D��Ŋ뜜�T�ށD�������Ɍ��ł��قƂ�ǎp�����邱�Ƃ��Ȃ��Ȃ����D������ł͋߂����̂Ǝv����D
�ʐ^�P�Q�D�I�X�͋��������ʂ܂ŐԂ��Ȃ�D9�����{����C�R�Ԃɂ�����O�̐��c�łӂ�ӂ�Ɣ��őŋ�Y������D
�ʐ^�P�R�D���Ă͊C�߂��̐����܂��w�Z�̃v�[���Ɏp�������Ă������C�ŋ߂͂��Ȃ�������ɂ��o�����Ă���D
�ʐ^�P�S�D���Ɍ��암�ł͐������������D�k���ł͂܂����Ȃ�̌̐������邱�Ƃ��ł���D
�ʐ^�P�T�D���n�̐����҂ŁC�r�̗������̎��n��̕����Ȃǂɂ�������D��ʂ������D
�ʐ^�P�U�D��ʂ������̂ł��̖�������D���n�̒����A���̔ɖ���r�ɏZ�ݒ����Ă���D���X�̊�ʂɔ�������o�邱�Ƃ�����D
�ʐ^�P�V�D��ʂɔ����������D���X�����ɂ́C�m�V�������o����̂Əo�Ȃ����̂�����D
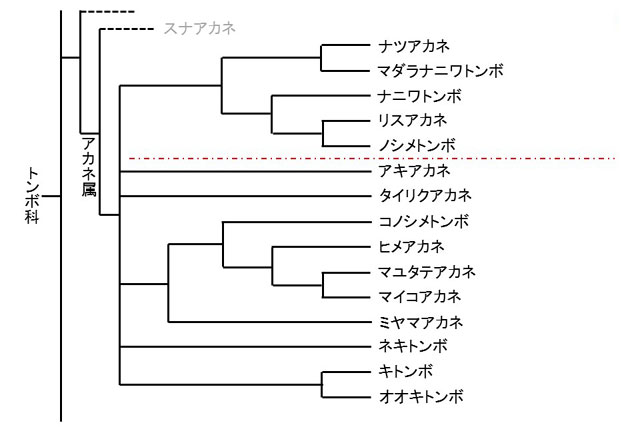
�@���āC������15��ɂ��āC�ŋ߂�DNA��p�����n�����ނ����Ă݂܂��傤�D�ȑO�̌`�Ԃ𒆐S�Ƃ������ނł͂�������Ƃ��Ȃ������C�Y�����@�Ɨc���̌`�Ԃ��C�����ɋ߉���̊Ԃň�v���邱�ƂɂȂ�܂����D���ɑŋ�Y�����s����Ɨc���̑����̒�����Q���P�n���ɂȂ��Ă���_�����ڂ���܂�
*1�D
�}�Q�D���Ɍ��ɐ�������A�J�l����DNA��͂Ɋ�Â��n������ (������C2012���Q��)�D
�@�Ԃ̈�_��������̒P�n���Q�͑ŋ�Y�����C�c���̑����������`���������Ă���D
*1.�ŋ�Y�����s���G�]�A�J�l�͂����5��Ƃ͒P�n�����Ȃ��Ȃ��D�܂������������Ȃ��D

��8���߂̑����̒������Ԃ̈�_�����̏�̌n���Q�Ɖ��̌n���Q�ňقȂ��Ă���D
�ʐ^�P�W�D�ŋ�Y������P�n���Q�̃A�J�l���g���{�D�c���̑�8���߂̑�������9���ߖ��[����D
�@������F�i�c�A�J�l�E�}�_���i�j���g���{�E�i�j���g���{�E���X�A�J�l�E�m�V���g���{�D
�ʐ^�P�X�D��L�ȊO�̌n���Q�̃A�J�l���g���{(�ꕔ)�D�Ő��E�œD�Y�����s���C�c���̑�8���߂̑�������9���ߖ��[���Ȃ��D
�@������F�A�L�A�J�l�E�^�C���N�A�J�l�E�R�m�V���g���{�E�}���^�e�A�J�l�E�I�I�L�g���{�D
�g���{�̐��Ԋw�@���m�O���t�F�A�J�g���{
�A�J�l���̐����j �P�D���Ɨc���̎���
�@���{�Y�̃A�J�l���̐����j�ɂ́C���Ɍ��Y�̃A�J�l���̉z�~�헪����l���āC�傫��2�̃^�C�v������ƍl�����܂��D����́C�c���z�~����^�C�v�Ɨ��z�~����^�C�v�ł��D����Ɏ��́C���z�~����^�C�v�ɂ́C2�̈قȂ�z�~�헪���Ƃ��Ă�����̂�����̂ł͂Ȃ����ƍl���Ă��܂��D����ł͂����ɂ��ďq�ׂĂ������Ƃɂ��܂��傤�D
�@�܂��c���z�~����^�C�v�ł��D���݁C�l�L�g���{�ƃ^�C���N�A�J�l�ɂ��āC�c���z�~���Ă���m���ȏ؋��������Ă��܂��D���́C�~�̏I����3��3��(2007�N)�ɁC�z�~�����l�L�g���{�̗c�����ʂɍ̏W�������Ƃ�����܂�(�����\)�D�l�L�g���{�̎������牺�ł̗����Ԃ́C���̃A�J�l���̎�Ɣ�ׂĔ��ɒZ���̂�������(�\�`�C�a�C�b�Q��)�C����̓A�J�l���ȊO�̑��̗c���z�~�ʂƂ���g���{��Ƃقڕς�肪����܂���D
�@�^�C���N�A�J�l�̕��́C���s�s���̊w�Z�̃v�[���ŗc���z�~���Ă��邱�Ƃ�����܂���(Matsura et, al., 1995)�D�^�C���N�A�J�l�͊w�Z�̃v�[���ő������������c���ŁC����ɂ���1���ɕ��ϓ�����1mm���x�̗c�����̂�Ă��܂��D����5��10��(1994�N)�Ƃ������������ɁC���{�s�̏��w�Z�̃v�[���ŁC��ʂ̃^�C���N�A�J�l�̏I��c�����̏W���Ă��܂�(�����\�G�E���O�ʐ^)�D����́CMatsura et al.(1995)�̎������z�~�c���̐����Ȑ�(p.294, Fig.2)�Ɉ�v������̂ŁC�c���z�~���Ă������Ƃ��������Ă��܂��D
�@�^�C���N�A�J�l�̎������牺�ł̗����Ԃ́C�\�`����33���ȉ��ƒZ�����̂������āC�����̐��l�͗c�����H�ɛz�����ėc���ʼnz�~����\�����������̂ł����C�t�ɁC100�����z���钷�����Ԃ̃f�[�^������܂��D���牺�ł͂��̂悤�ȕϓ��͌��\����ɂ��Ă��C���Ȃ��Ƃ������Ԃ������ԂɂȂ鉽�炩�̏����͑��݂���ƍl�����C���z�~����̂̑��݉\����ے肷�邱�Ƃ͂ł��Ȃ��ł��傤(cf. �V���E�ߓ��C1989)�D
�@Ando(1967)�̓g���{�ڂ����ߒ��̏ڍׂȌ������ʂ���C���g�̌����ɗp�������{�Y�̃g���{��21��̗����C�x�����̂Ȃ�����(������7-40��*2)�ƁC�x�����̂��闑(������81-231��)�ɕ����Ă��܂��D����͒P�ɗ����Ԃ���K���ɕ��������̂ł͂Ȃ��C���������̉ߒ������Ȃ���o�������_�ł��̂ŁC�x�����Ɋւ��Ă͐M���ł��鐔�l�Ƃ����܂��D���̐��l�������ɂ���ƁC�^�C���N�A�J�l��33���ȉ��̃f�[�^�͔�x�����C137���C150���Ƃ����̂͋x�����̗��ƂȂ�܂��D���ꂩ��l����ƁC�^�C���N�A�J�l�̗��́C�����ɂ���ċx��*3�����肵�Ȃ������肷�闑�ł��邱�Ƃ���������܂��D
�@Corbet(1962)�́CAndo�̃g���{��������(�Ⴆ��Ando�C1957)����b�ɂ��āC�����Ԃ���r�I�Z������ direct development egg (����邱�ƂȂ��������闑�Ƃ����Ӗ�)�C��r�I�������� delayed development egg (�x���������������闑)�ƌĂ�ł��܂��D�u�x��*4�v�Ƃ����������ɔ����Ă��銴�������܂����C�u�x�������Ȃ��������闑�v�ƍl���č��������Ȃ��Ǝv���܂��D
�@�ł͎��ɏ�L2��ȊO�ɂ��čl�@�������Ă݂܂��傤�D�����2��ȊO�̃A�J�l���̎��炳�ꂽ�����Ԃ�����ƁC������[�����ƂɋC�Â��܂��D���Ɍ��암�ɐ������Ă���A�J�l���̂����C���X�A�J�l�C�m�V���g���{�C�}�_���i�j���g���{�C�i�j���g���{�̗����ԃf�[�^�ɁC��r�I�Z���Ԃ̂��̂������܂܂�Ă��邱�Ƃł�(�\�a)�D�����̃A�J�l���̃g���{�͂��ׂđŋ�Y��������g���{�ł����āC���𐅂̂Ȃ����y��ɗ��Ƃ���ނ���ł��D���́C�ŋ�Y�������Ȃ����z�~�A�J�l���ł́C�T��100�����z����f�[�^�������Ȃ��Ă��܂�(�\�b)�D
�@�ߓ�(1994)�ł͓~�G�ɉ��O�ɒu���Ď��炵���m�V���g���{�C�i�j���g���{�̍ŒZ�����Ԃ̃f�[�^�������āC���ꂼ��138���C148���Ǝ�������Ɣ�ׂĔ��ɒ����Ȃ��Ă��܂�(�\�a)�D����́C�����炭�ቷ�ɂ���Ĕ��炪��~�������Ԃ����邽�߂ł��傤�D�܂��C�����4��̏ꍇ�ł��C�����炭��������̈Ⴂ�ɂ���āC�����Ԃ��ʏ�̗��z�~����݂�100�����z����ꍇ�����邱�Ƃ�������܂�(�\�a)�D����ɁC�����ŋ�Y�����s���i�c�A�J�l�̏ꍇ�ɂ́C�������������Ă������Ԃ��ɒ[�ɒZ���Ȃ邱�Ƃ͂Ȃ��Ƃ������ʂ��o�Ă��܂�(�\�a)�D
�@��c(1996)�́C�ŋ�Y�����s����ɂ��ė������Z���X�������邱�Ƃ��w�E���Ă���CCorbet(1962)�̋c�_�����p���C�u���ɐZ��������Ԃł͗��̋x���������Ȃ����C���邢�͒Z���Ȃ��Ă��܂���́C���z�~���ێ����邽�߂ɂ͗��𐅂̂Ȃ��Ƃ���ɒu���K�v������C���̂Ȃ��ꏊ�ɗ����Y�ނ��Ƃ��\�ɂ���ŋ�Y���́C���̂悤�Ȑ����̗�����������ɂ����ėL���ȎY�������ł���ɈႢ�Ȃ��v�ƈ�̍l�������q�ׂĂ��܂��D�Ȃ��C�i�c�A�J�l�́C�ŋ�Y�����s���ɂ�������炸�C�����Ԃ��T��100�����z���Ă���C�x�����̗��ł���\��������܂��D
�@���R��Ԃł́C�ŋ�Y�������Ă����̗����K���������ɐZ����Ȃ��ۏ͂���܂���D���Ƀ��X�A�J�l��7������Y�����Ă���̂��ώ@���Ă��܂�(�ʐ^�R�O�E)�̂ŁC���ɐZ����C���̍����Ăɛz�����邱�Ƃ͂��蓾�邱�Ƃł��傤�D���������āC���X�A�J�l���c���œ~���}����\���͏\���ɂ���Ƃ����܂��D�i�j���g���{�C�m�V���g���{�C�}�_���i�j���g���{�́C���̊ώ@�ɂ���9���ȍ~�ɎY�����Ă��܂����C�g�����H�ł����͂�N���ɛz������\���͂Ȃ��Ƃ͂����܂���D�ȏ�̂��Ƃ���C�ŋ�Y�����s����ɂ��ẮC�c���œ~���}���Ă���\��������Ƃ����܂��D�������C�c���œ~���z���邩�ǂ����͕�����܂���D
�\�`�D���Ɍ��암�n��ɐ�������c���z�~���A�J�l���́C��Ɏ�������ɂ�闑����
*1
|
| �l�L�g���{ |
8�� |
��2 |
|
8�� |
��2 |
|
11�� |
��3 |
|
14�� |
��2 |
|
15�� |
��1 |
|
16�� |
��3 |
|
| �^�C���N�A�J�l |
15�� |
��3 |
|
30�� |
��1 |
|
33�� |
��4 |
|
137�� |
��4 |
|
150�� |
��1 |
��1. ���g���{�k�b��(1984)
��2. �V���E�ߓ�(1989)
��3. �ߓ�(1994)
��4. ����(1967)
|
*1. �̗�����Ă���z������܂ł̊��ԁD�����̗��Ŗ��L����ꍇ�͍ŒZ�̂��̂��̗p���Ă���

5��10���ɐ��{�s�̏��w�Z�v�[���ō̏W���ꂽ�^�C���N�A�J�l�̏I��c���D
*2. Corbet (1999)�́C���̌�̌������܂߂āC�x�����̂Ȃ��C���ڔ������闑�̎Y������Ӊ��܂ł̓�����5-60���Ƃ��Ă���D
*3�D�����͋x�~(quiescence)�Ƃ��������K��������Ȃ����C�L�`�̋x���Ƃ����Ӗ��Ŏg�p�����D
*4�D�{���͋x�����Ă��邱�Ƃ̏ؖ��͓���Ǝv����̂Ŕ���x���܂��͋x�~�Ə����ׂ��ł��낤���C�����ł͍L�`�̋x���̈Ӗ��Ŏg�p�����D
�\�a�D���Ɍ��암�n��ɐ�������ŋ�Y�����s���A�J�l���́C��Ɏ�������ɂ�闑����
*5
|
| ���X�A�J�l |
41�� |
��1 |
|
43�� |
��2 |
|
69�� |
��2 |
|
74�� |
��2 |
|
94�� |
��2 |
|
| �m�V���g���{ |
15�� |
��3 |
|
30�� |
��1 |
|
46�� |
��2 |
|
106�� |
��2 |
|
148�� |
��2 |
| �@�i���O����j |
138�� |
��3 |
|
| �i�j���g���{ |
28�� |
��4 |
|
35�� |
��1 |
|
93�� |
��2 |
|
105�� |
��2 |
|
140�� |
��2 |
|
223�� |
��2 |
| �@�i���O����j |
148�� |
��2 |
|
| �}�_���i�j���g |
28�� |
��2 |
| ���{ |
30�� |
��4 |
|
35�� |
��1 |
|
41�� |
��2 |
|
58�� |
��2 |
|
| �i�c�A�J�l |
96�� |
��2 |
|
110�� |
��2 |
|
161�� |
��2 |
|
166�� |
��1 |
��1. ���g���{�k�b��(1984)
��2. �V���E�ߓ�(1989)
��3. �ߓ�(1994)
��4. �ߓ�(1995)
|
*5. �̗�����Ă���z������܂ł̊��ԁD�����̗��Ŗ��L����ꍇ�͍ŒZ�̂��̂��̗p���Ă���D����͗��𐅂ɐZ������Ԃōs���Ă���Ǝv����D

�ʐ^�Q�O�D�Ő��Y��������R�m�V���g���{(��)�̗����Ԃ�100���C�ŋ�Y�������郊�X�A�J�l(�E)�̗����Ԃ�41��(���g���{�k�b��C1984)�D
�@�ł́C�\�`�C�a�Ɍf����7��ȊO�̃A�J�l���ɂ��čl���Ă������Ƃɂ��܂��傤�D�����͂��ׂāC�Ő��Y���܂��͑œD�Y�����s����ł��D
�@Ando(1962)�́C�u�~�����̗��ɂ�3�̃^�C�v�������āC�����}�^�C�A�I�C�g�g���{�^�C�A�J�l�^�ɕ�������D�A�J�l�^���������������i�K(
full grown stage)�œ~������v�Əq�ׂĂ��܂��DAndo(1962)�Ŏg��ꂽ�ޗ��̒��ŃA�J�l���̓A�L�A�J�l�ł����D�܂����������ɂ��ďڍׂɊώ@���Ă����n��(1996)�́C�u�A�L�A�J�l�̗��͎������炷��ƁC���]���O��������قڊ��������i�K�Ŋ�������߂����D�����ł͈ꌎ����O�����炢�܂Ŏv���o�����悤�ɛz�����������C�����Ȃ���Λz���ł��Ȃ��v�Əq�ׂĂ��܂��D�܂���c(1993)�́C�A�L�A�J�l�̋x���̃X�e�[�W�͎��牷�x�ɂ���đ傫���ω�����������Љ�Ă��܂��D���̒���18���ł̎���̏ꍇ�C�u��(��_)����ł��������āC�t�������قƂ�ǂł�����������Ԃ܂�20���Ԃقǂő��₩�ɒB���C......�C���̒i�K�ŃX�g�b�v���Ă��܂��C......�C100���ڂ��炢�ɂȂ��Ă悤�₭�z�����n�܂�܂��v�ƋL�q���Ă��܂��D
�@�����̂��Ƃ���l����ƁC�A�L�A�J�l�́C���R��Ԃł́C�H�ɂق���������������ԁC����������ƛz�����O�̏�Ԃ܂Ŕ������������C���̒i�K�ŋx���ɓ���ƍl�����܂��D�����āC�~�̊Ԃɋx��������I���C�x�����Ŕj����C�t�̋C���㏸�Ɛ��̑��݂ɂ���ěz��������̂ƍl�����܂��D
�@�A�L�A�J�l�ȊO�̃A�J�l���̎킪��L�Ɠ����Ƃ����邩�ǂ����͕�����܂���D�\�b���������C�����Ԃ́C���Ȃ��Ƃ�50�����z����C��r�I�������̂������Ȃ��Ă��܂�(�I�I�L�g���{��29���C50���ƒZ�����̂�����܂����C�����͈��Q���Y�̂��̂ł�)�D�ł�����C�\�b�̎�ɂ͉��炩�̋x������������Ă���ƍl���Ă�������������܂���D�����̎�̗��͐��ɐG���ꏊ�ɒu����C�����������~�̌����ɂȂ�ɂ������Ƃ���C�����̉z�~�헪�͎��̂悤�ɉ����I�ɂ܂Ƃ߂邱�Ƃ��ł���ł��傤�D
- �H�ɎY�����s����
- �Y�����ꂽ���������n�߂�D
- ���̓���̒i�K�ŋx���ɓ���D
- �~�̊Ԃɋx�����Ŕj����C�ቷ�ɂ�锭���~���(�x�~)�ɂȂ�D
- �������㏸���C�����_�̉��x���z�����Ƃ��ɔ�����ĊJ����D
- �����ěz���Ɏ���D
�@���̋x���̑��̈Ӌ`�́C�ϊ����̂��闑�ň��S�ɓ~�����邱�Ƃɂ���ƍl�����܂����C������d�v�ȈӖ�������܂��D����͂��̌�̐����j�̓��������m�ۂ��邱�Ƃł��D9�����납��n�܂�11������܂ő����A�J�l���̎Y���V�[�Y���ɂ͂��Ȃ蕝������܂��D�������C�x���ɂ�����̒i�K�ł̔����~�́C�x���Y���������̌�ꂽ�������̐i��ł��闑�ɒǂ������Ƃ��\�ɂ��Ă��܂��D����ɓ~�̊��������ɋx�����Ŕj�����ƁC�����x�������_��臒l���z�����t�ɁC�����i�K�Ŕ����~���Ă���������Ăɔ�����ĊJ����̂ŁC���̌�̛z���������I�ɋN����ƍl������̂ł��D�ł�����C���ꐶ���ꏊ�ł̉H�����ꎞ���ɏW�����邱�Ƃɂ��Ȃ����Ă����܂��D
�@�̌Q�ɂ���ăA�J�l���c���̏o���������قȂ�̂́C���̔����_���z���鉷�x�̎��������^�C�~���O���C�Ⴆ�C���̒n�̕W���C�ܓx�C���邢�͓��A���������Ƃ������悤�ȋǏ��I�����Ȃǂɂ���Ĉ���Ă��邩�炾�ƍl����悢�ł��傤�D�܂��C�L�����z�������ł́C�n��̌Q�ɂ���āC�����_�̉��x���قȂ��Ă��邩������܂���D
�@�ȏ�C���̎���ɂ��čl�@���Ă��܂����D�m���Ȃ��Ƃ͂����Ȃ��ɂ��Ă��C���ǂ̂Ƃ���C�l�L�g���{�����͕ʂɂ��āC�u(���Ɍ��Y��)�A�J�l���ɂ����ẮC�������ʂ̉z�~�X�e�[�W�ŁC�����ɂ���ėc���z�~����̂����݂���������v�Ƃ������Ƃ͊m���ɂ�����Ǝv���܂��D�������C�c���z�~�Ɋւ��ẮC�~�G�ɂ��̗c�������ł���\�������邱�Ƃ́C�t�������Ă����K�v������ł��傤�D
�@�ł́C�����Ď��̃X�e�[�W�C�c������ɂ��čl���Ă݂܂��傤�D�A�J�l���̗c���́C��ʂɂ��Ȃ葬����������ƍl�����Ă��܂��D�������c���̎���L�^������ƁC���ɔ��ɒ������Ă���ꍇ�����邱�ƂɋC�Â��܂��D�Ⴆ�C�}���^�e�A�J�l218���C���X�A�J�l230���C�A�L�A�J�l195���C�l�L�g���{306��(����������g���{�k�b��C1984)�ȂǁC�ق�200�����z��������ɂȂ��Ă��܂��D�g���{�c���̐������x�́C���x��a�ȂǁC��������ɑ傫�����E����܂��D���̎����ɂ͎��玞�̏�������Ă��܂���̂ŁC�����c�����Ԃ̐��l�������Ė�O�̏�Ԃf���邱�Ƃ͉ߌ�������Ǝv���܂��D
�@�����ŁC������H���܂Ŏ�����p�������L�^���W�߂āC�g�[�^���ʼnH���������ɋN���邩���܂Ƃ߂��̂��\�c�ł��D������s���������Ƃ͎��̒m�l�ŁC����ɂ͐��ʂ��Ă���C���̃f�[�^�͐M������������̂��Ɣ��f�ł��܂��D���������ƁC�������������Ɋւ�炸�C�H���́C��O�Ȃ��̗����N�̏t���珉�Ăɂ����ċN���Ă��邱�Ƃ�������܂��D���ꂪ�Ӗ����邱�Ƃ́C���R�����ŗ��z�~�C�����ėc���z�~���Ă����Ƃ��Ă��C�����j�̘g�g�݂Ƃ��āC��N�ꉻ�ŁC���H�����č��܂łɍs����悤�Ȃ����݂��A�J�l���̑����̎�ɔ�����Ă���Ƃ������Ƃł��D���̂��Ƃ͎��ۂ̖�O�ώ@�ɂ��C���ɂ悭���v���Ă��܂�(�ʐ^�Q�P)�D
�@�ȏ�̂��Ƃ��܂Ƃ߂܂��ƁC��ʓI�ɁC���z�~�����t�ɛz�������c���́C�}���ɐ������C���Ă܂łɗc���Ƃ��Ă̐������������ĉH������C�����āC�c���z�~�����ꍇ���C�����_�������Ɨc���͋}���ɐ������ĉH������C�Ƃ����Ă悢�ƍl�����܂��D�������C��O�I�ɒx���H������ꍇ�����邱�Ƃ͕t�������Ă����܂��傤�D�L�g���{�ł�9��23���ɉH������̌̂��ώ@���Ă��܂�(�ʐ^�Q�V�Q��)�D
�@���āC�c������̍Ō�̃C�x���g���C���̍ŏI�̒E��C�H���ł��D�A�J�l���̉H���͗�O�Ȃ��|���^�ł��D���������钆�ɉH�����܂�(�ʐ^�Q�Q)�D6���̒����{�̑����ɐ����n�֍s���Ă݂�ƁC���ۂ̐A�����珈����s�ɔ�ї��A�J�l���̃g���{���ώ@���邱�Ƃ��ł��܂��D���Ƀ^�C���N�A�J�l��I�I�L�g���{�̂悤�ȁC���ɒW���F�����Ă���g���{�́C�H������ɂ������Z���F�Â��Ă��Ĕ��������̂ł�(�ʐ^�Q�R)�D
�\�b�D���Ɍ��암�n��ɐ������邻�̑��̃A�J�l���́C��Ɏ�������ɂ�闑����
*6
|
| �A�L�A�J�l |
72�� |
��3 |
|
76�� |
��2 |
|
85�� |
��2 |
|
103�� |
��2 |
|
106�� |
��2 |
|
111�� |
��2 |
|
127�� |
��1 |
|
| �R�m�V���g���{ |
100�� |
��1 |
| �@�i���O����j |
131�� |
��3 |
|
| �q���A�J�l |
109�� |
��2 |
|
136�� |
��2 |
|
153�� |
��2 |
|
161�� |
��1 |
|
171�� |
��2 |
|
| �}���^�e�A�J�l |
54�� |
��1 |
|
58�� |
��3 |
|
64�� |
��3 |
|
88�� |
��2 |
|
105�� |
��2 |
|
112�� |
��2 |
|
116�� |
��2 |
|
138�� |
��2 |
|
| �}�C�R�A�J�l |
51�� |
��1 |
|
58�� |
��2 |
|
61�� |
��2 |
|
67�� |
��2 |
|
107�� |
��1 |
| �@�i���O����j |
140�� |
��3 |
|
| �~���}�A�J�l |
191�� |
��1 |
| �@�i���O����j |
130�� |
��3 |
|
| �L�g���{ |
106�� |
��2 |
|
121�� |
��2 |
|
134�� |
��1 |
|
| �I�I�L�g���{ |
29�� |
��4 |
|
50�� |
��4 |
|
106�� |
��2 |
|
108�� |
��2 |
|
123�� |
��2 |
|
136�� |
��2 |
|
168�� |
��1 |
|
186�� |
��1 |
| �@�i���O����j |
158�� |
��3 |
��1. ���g���{�k�b��(1984)
��2. �V���E�ߓ�(1989)
��3. �ߓ�(1994)
��4. �ߓ�(1995)
|
*6. �̗�����Ă���z������܂ł̊��ԁD�����̗��Ŗ��L����ꍇ�͍ŒZ�̂��̂��̗p���Ă���
�\�c�D���Ɍ��암�n��ɐ�������A�J�l���̗̍�����H���܂ł̎���L�^
*7
|
| �̗��� |
�H���� |
|
|
| �i�c�A�J�l |
|
|
| 10/-8 |
7/18 |
��1 |
|
| �}�_���i�j���g���{ |
| 9/15 |
7/18 |
��21 |
|
| �i�j���g���{ |
| 9/-4 |
6/21 |
��1 |
| 10/-9 |
7/19 |
��1 |
|
| ���X�A�J�l |
| 9/-4 |
5/24 |
��1 |
|
| �m�V���g���{ |
| 10/-8 |
4/29 |
��1 |
|
| �A�L�A�J�l |
|
|
| 10/21 |
6/-4 |
��2 |
|
| �^�C���N�A�J�l |
|
|
| 10/20 |
6/10 |
��2 |
|
| �R�m�V���g���{ |
| 10/23 |
7/25 |
��2 |
|
| �}���^�e�A�J�l |
| 8/24 |
6/-1 |
��1 |
| 9/15 |
7/28 |
��1 |
| 9/-1 |
4/19 |
��2 |
|
| �}�C�R�A�J�l |
| 10/-8 |
6/23 |
��1 |
|
| �~���}�A�J�l |
| 11/17 |
7/23 |
��2 |
|
| �l�L�g���{ |
| 9/22 |
7/-2 |
��2 |
| 8/-3 |
3/-7 |
��2 |
|
| �I�I�L�g���{ |
| 10/-9 |
7/20 |
��1 |
| 10/20 |
8/13 |
��2 |
| 10/20 |
8/23 |
��2 |
��1. �V���E�ߓ�(1989)
��2. �ߓ�(1994)
|
*7. �H���͂��ׂč̗��̗��N�ł���̂ŁC���t�������L�ڂ����D

�ʐ^�Q�P�D���F���X�A�J�l�̏I��c���D2011.5.28.�@�E�F���X�A�J�l�̉H���D2007.6.10.�@
�@�o���I�ɏI��c�����̂�鎞�����H���������鎞���������������܂��Ă��邱�Ƃ́C�����j�����܂��R���g���[������Ă��邱�Ƃ̏ł���D
�ʐ^�Q�Q�D���F�A�L�A�J�l�̉H���C�|���^�̉H��(�Î~�p��)�C2011.6.19.�C�E�F�l�L�g���{�̉H���k�C��O�C2012.6.23.
�ʐ^�Q�R�D���F�^�C���N�A�J�l�I�X�̉H����Ԃ��Ȃ��́D2012.6.10.�@�E�F�I�I�L�g���{���X�̉H������̌́D2018.6.28.
�g���{�̐��Ԋw�@���m�O���t�F�A�J�g���{
�A�J�l���̐����j �Q�D�����̎���
�@�ł͎��̃X�e�[�W�C�����ɂ��čl���Ă������Ƃɂ��܂��傤�D���ׂẴg���{��ɂ́C��Ȃ菬�Ȃ萫�I�ɐ��n����܂ł̖��n����(�O���B���ԂƂ����܂�)�����݂��܂��D�����g�̊ώ@����C�x�����̂Ȃ��g���{�ł́C���̒����͐�������2�T�Ԓ��x�ł���Ƃ����܂��D��ʓI�ɁC�l�L�g���{�������A�J�l���ł́C6�C7�����炢�ɂ͉H�����H�ɔɐB�������J�n����܂ł����I�ɖ��n�Ȋ��Ԃƍl�����܂�����C���̒����́C���Ȃ��Ƃ�1�����ȏ�C�������̂ł�3�������x�͂���ƍl�����܂��D
�@��������������x���͈�ʂɉċx��(summer diapause)�ƌĂ�Ă��܂�(Masaki, 1980)�D�x���Ƃ����Ă��������~����킯�ł͂���܂���D��c(1993)�́C�A�L�A�J�l�ɂ��āC���̊ԗ����̐��n���}������Ă���Ƃ������ƂŁu���B�x���v�Ƃ�������g���Ă��܂��D���̐��B�x�����C���ɂ���ėU�����ꉽ�ɂ���đŔj����邩�Ƃ��������́C�قƂ�NJF���ł��D�������C�A�L�A�J�l�ɂ��āC�H�������ƔɐB�����J�n�����C�����Ă���܂ł̑O���B���Ԃ̒����ɁC�n���I���z(�N���C��)������炵�����Ƃ��C��c(1993)�͎w�E���Ă��܂��D�܂�C���g�Ȓn��قljH�������������Ȃ�C�ɐB�����J�n�������x���Ȃ�C���̌��ʑO���B���Ԃ������Ȃ�Ƃ������Ƃł��D
�@�O���B���Ԃ��������Ƃɂ��ẮC���ۂɖ�O�Ő������������Ă���قƂ�Nj^���̂Ȃ������Ƃ��Ď���邱�Ƃ��ł���ł��傤�D�����C���̗��R�₵���݂��悭������Ȃ��Ƃ������Ƃł��D���������������ł悭���������ɏo�����̂��C�A�L�A�J�l�͂Ȃ����̑O���B���Ԃɍ����R�ɏオ��̂��C�Ƃ������₢�����ł��D�ڍׂȊώ@�����ƂɃA�L�A�J�l�̈ړ��m�ɂ����̂́C�n��(1953)���낤�Ǝv���܂��D���̌セ�̗��R�ɂ��Ă��낢��ȍl�����\������܂����D�����͏�c(1993)�ɑ�������Ă��܂��̂ŁC�����̂�����͂�������������������D

�ʐ^�Q�S�D���F�H���セ�̂܂ܐ��c�̈�̗t�Ɏ~�܂��Ă���A�L�A�J�l�D2016.6.24.�C�E�F�R���t�߂̕����ʼnĉz�������Ă���A�L�A�J�l�D2010.8.14.
�@�ł̓A�L�A�J�l�ȊO�̃A�J�l���́C�O���B�����ǂ��ʼn߂����Ă���̂ł��傤���D�}���^�e�A�J�l�C�q���A�J�l�C�}�C�R�A�J�l�C�~���}�A�J�l�C�}�_���i�j���g���{�Ȃǂ́C��������߂��̎��ѓ��C�щ��C�����ȂǂŐ������Ă��܂��D�i�c�A�J�l�͎R�̘[������ɏW�c�ł���Ƃ�����ώ@�������Ƃ�����C�^�C���N�A�J�l�́C8���ɁC�R�̗Ő��𑖂铹�H��ŁC���ォ��~��Ă����̂��������Ƃ�����܂��D�R�m�V���g���{�͎R�[�̎��тłƂ��ǂ��������܂��D�m�V���g���{�͎R���t�߂̑����Ō������Ƃ�����܂��D�O���B���̌̂������ɂ����̂��C�L�g���{�C�I�I�L�g���{�ł��D�H������̌̂��������Ƃ͂���܂����C���̌�ǂ��Ő������Ă���̂��C���͂����̖��n�̂������������Ƃ�����܂���D������(1999)�ɂ��ƁC���т���ɂ���Ə�����Ă��܂��D
�ʐ^�Q�T�D���F�}�C�R�A�J�l�̑O���B���̃I�X�D2016.8.3.�C�E�F�q���A�J�l�̑O���B���̃I�X�D2016.8.3.�C
�@�Ƃ��ɉH������߂��̗щ��őO���B�����߂����D
�ʐ^�Q�U�D���F�ѓ��̔��Â��Ƃ���ŋx��ł���O���B���̃R�m�V���g���{�I�X�D2013.8.18.�C
�@�E�F�щ��̑��n�ŋx�ޑO���B���̃~���}�A�J�l���X�D2010.7.31.
�ʐ^�Q�V�D���F�I�I�L�g���{�̉H�������̃��X�D2009.6.21.�C�E�F�L�g���{�̉H����Ԃ��Ȃ������̃I�X�D2006.9.23.�C
�@�Ƃ��ɉH����̑O���B���̌̂�������̂�����D�L�g���{�͂��Ȃ�x�������ɉH��������̂�����C���~�܂Ŋ������Ă���D
�@���̂悤�ɁC�����̃A�J�l���̎�̑O���B���Ԃ͒����Ȃ��Ă��܂����C��r�I�Z���ƍl�����������܂��D�܂��͐��B�x�����s��Ȃ��ƍl������̂��C�l�L�g���{�ł��D�l�L�g���{��Corbet(1962)�̒�`����ċG��(summer species)*1�Ƃ����鐶���j��L���Ă��܂��D�܂�C�l�X�̗�ʼnz�~�����c�������X�ɉH�����C�H��������Z���O���B���Ԃ��߂��������ƔɐB�������s���܂��D���������āC�ɐB�������s�������̏o�����Ԃ͂��Ȃ蒷���Ȃ�܂��D5������Y�����Ă���(������C2012)�Ƃ����L�q������C����ɋߓ�(1994)��R�{��(2009)�́C�l�L�g���{����N���Ă���\�����w�E���Ă��܂��D����6��17���ɎY�����Ă���l�L�g���{��10��18���Ɍ�����Ă���y�A(�ʐ^�Q�W)���C�܂�8��24���ɐ��n�����I�X��H�����ĊԂ��Ȃ����n�ȃ��X���L�^���Ă��܂�(�ʐ^�Q�X)�D
*1. Corbet(1962)�̓g���{�̐����j�̘g�g�݂��Csping species �� summer species �ɕ����ďЉ�Ă���Dspring species �Ƃ́C�ʏ�C�~���I��c���ʼn߂������̂��������N�̏t�ɉH�����ĔɐB�������s���悤�Ȏ�������Csummer species �Ƃ́C�l�X�̗�ʼnz�~�����c�������̊��Ԏ��X�ɉH�����ĔɐB�������s���悤�Ȏ�̂��Ƃ������D
�ʐ^�Q�W�D�l�L�g���{�́C���Ɍ��ł��C6������10���܂ŔɐB�����𑱂��Ă���ċG��ł���D
�@���F6��17��(2012�N)�ɎY�����s���Ă���l�L�g���{�C�E�F10��18��(2009�N)�Ɍ�����Ă���l�L�g���{�D
�ʐ^�Q�X�D���F�l�L�g���{�̐��n�����I�X���̂Ă���Ɏ~�܂��Ă���D2009.8.24.�C
�@�E�F�������Ƀl�L�g���{�̖��n�ȃ��X���r�̔ȂɎ~�܂��Ă���D2009.8.24.
�@���ɒZ���ƍl������̂́C���X�A�J�l�ƃi�j���g���{�ł��D�����͂�������ŋ�Y�������Ă����ŁC�c���z�~�̉\�����w�E���܂����D���X�A�J�l�ƃi�j���g���{�́C���n�Ȏ����ɂ͐�������߂��̎��ѓ��ɂ��܂��D7���ɂ̓i�j���g���{�ł͐D�F�̕��𐁂��Ă���C���ł�7��7���Ƀ^���f���́C7��15���ɎY�����ώ@����Ă��܂�(�{��C2018)�D�܂����X�A�J�l�ł�7�����{�ɂ͕������Ԃ��Ȃ��Ă��āC�Y�����Ă���̂��ώ@���Ă��܂�(�ʐ^�R�O)�D���Ɍ��암�ɂ�����C���̃��X�A�J�l�Y���̍ł������ώ@�L�^�́C7��21��(2008�N)�Ƃ������̂ł��D
�ʐ^�R�O�D���F7��24���ɂ����D�F�̕��𐁂��Ă���i�j���g���{�̃I�X�D2011.7.24.�C
�@�E�F7��31���ɔ��Â����ѓ��ŎY�������Ă��郊�X�A�J�l�D2016.7.31.
�@���̂悤�ɁC�O���B���Ԃ̒�����ɐB�������n�߂鎞���ɂ͂��Ȃ�����������̂́C�u�A�J�l���̃g���{�͏H�ɔɐB�������s���Ă���v�Ƃ����̂́C�ԈႢ�Ȃ������Ƃ����܂��傤�D�ł́C�����̃g���{�͂��܂ŔɐB�������s���Ă���̂ł��傤���D���x�͂��̏I���̎����ɂ��āC�ώ@��L�^�����Ƃɍl���Ă݂܂��傤�D�����I���̎����͋C���̈قȂ���{�e�n�ň���Ă���Ǝv���܂��̂ŁC�����ł͕��Ɍ��𒆐S�Ƃ����n��Ɍ��肵�čl���邱�Ƃɂ��܂��D
�@�܂������Ƃ��x���ƌ����Ă��悢�̂́C�L�g���{�ł��傤�D�L�g���{�͔N���z�����Ƃ��Ƃ�����̂ŗL���ł��D�R�{��(2009)�ɂ́C1��9���ɕ��Ɍ��Ō���C�Y�����ώ@�C1��22���ɃI�X2����ڌ������Ƃ����L�ڂ�����܂��D����1��8��(2019�N)�ɔɐB�������ώ@���Ă���C���ꂪ�����Ƃ��x���L�^�ł��D���̔N�C�L�g���{��1��13���܂Ő����c���Ă���̂��m�F���܂����D
�ʐ^�R�P�D���F1��8���ɎY�������Ă���L�g���{�̃y�A�D2019.1.8.�C�E�F1��13���ɂ܂��r�ȂŊ��������Ă���L�g���{�̃I�X�D2019.1.13.
�@�L�g���{�ȊO�̃A�J�l���̃g���{�ɂ��ẮC���̊ώ@�L�^�𒆐S�ɁC�ꕔ��(2010)�̋L�^���Q�Ƃ������̂���C�x���L�^��\�d�ɏЉ�܂��D�����̃f�[�^�́C�K�����������܂Ŋ������Ă��邩��ړI�Ƃ����������ʂł͂Ȃ��̂ŁC����ȍ~�ɂ��ɐB�������s���Ă�����C�������Ă��邱�Ƃ͏\�����蓾�܂��D���̂悤�Ȍ��E�͂�����̂́C�\�d����C�A�J�l���̃g���{�́C��������10�������ς��܂ł͊e��Ƃ��ɐB�������s���Ă���C�ꕔ�̎��11������12����{�܂ŔɐB�������s�����Ƃ�����C�ƌ��_�Â��Ă��悢�Ǝv���܂��D���̃f�[�^������C��͂�L�g���{�̊����̒x�����ۗ����Č�����Ǝv���܂��D
�@�ɐB�������I������̐������Ԃ��C�㐶�B�� post reproductive period �ƌĂт܂��D����ɂ��ẮC���̌̂̒u���ꂽ��C���̔N�̋C�ۏȂǂ̉e���ɂ���Č��܂��Ă��܂����C���Ɍ��ł́C�^�C���N�A�J�l��2020�N�ɔN���z�����̂��m�F���܂������C�قƂ�ǂ̏ꍇ�C�L�g���{�ȊO�͔N���z�����Ƃ͂Ȃ��Ƃ�����ł��傤�D
�ʐ^�R�Q�D�x�������̃A�J�l���̎Y���D
�@�㍶�F�R�m�V���g���{�C2013.12.5.�C��E�F�A�L�A�J�l�C2011.12.4.�C
�@�����F���X�A�J�l�C2012.11.25.�C���E�F�}���^�e�A�J�l�C2011.12.4.
�\�d�D���Ɍ��암�n��ɂ�����A�J�l���̒x���L�^(10���ȍ~�̕��̂�)�D
|
| �ɐB���� |
�����m�F |
|
| �i�c�A�J�l |
| 11/20('11) |
*11/24('02) |
| �}�_���i�j���g���{ |
| 10/13('12) |
*11/10('05) |
| �i�j���g���{ |
| 10/10('10) |
*11/16('01) |
| ���X�A�J�l |
| 11/25('12) |
12/-4('11) |
| �m�V���g���{ |
| 10/10('10) |
11/20('11) |
| �A�L�A�J�l |
| 12/-4('11) |
12/25('18) |
| �^�C���N�A�J�l |
| 11/18('18) |
01/01('20) |
| �R�m�V���g���{ |
| 12/-5('13) |
12/16('25) |
| �q���A�J�l |
| 10/-9('11) |
12/-6('08) |
| �}���^�e�A�J�l |
| 12/-4('11) |
12/12('10) |
| �}�C�R�A�J�l |
| 10/10('10) |
*11/24('02) |
| �~���}�A�J�l |
| 10/-6('13) |
*11/12('00) |
| �l�L�g���{ |
| 10/13('12) |
11/15('18) |
| �I�I�L�g���{ |
| 11/18('18) |
**12/-9('25) |
|
*��͓�(2010)�ɂ��L�^�D����ȊO�͂��ׂĎ��g�ɂ��ώ@�L�^�D����������Ɍ��̋L�^�Ɍ���D('01)�Ȃǂ͐���N��\���D
**�I�I�L�g���{�ɂ��ẮC12��25���ɕ��Ɍ��Ŗڌ����ꂽ���Ƃ��f���ɓ��e���ꂽ�D�ʐ^�����Ȃ��̂Œ��ɂƂǂ߂��D
�g���{�̐��Ԋw�@���m�O���t�F�A�J�g���{
�A�J�l���̐������C���ɎY���ꏊ�Ɨc���̐�����
�@�A�J�l���̃g���{�́C���̂قƂ�ǂ��~�����̃g���{�ł��D�B��̗�O�Ƃ�����̂́C�~���}�A�J�l�ł��傤�D�~���}�A�J�l�̗c���̔��������ꏊ�ɂ��ẮC��ɐ�������̌Q���g���ďڂ������ׂ��܂���(Higasikawa et al., 2016)�D����ɂ��ƁC3�|4���ɛz������������1mm�ȉ��̗c���͐�̒��ł����̗���Ă��Ȃ������ɑ��������C��������ɂ�Ď���ɗ���̑����ꏊ�Ɉړ����Ă����C�Ƃ������̂ł��D���������āC�~���}�A�J�l���������邽�߂ɂ́C�~�������Ɨ��������C�����Ă��̈ڍs�������A���I�ɂȂ����Ă���悤�Ȑ������K�v���Əq�ׂĂ��܂��D�~���}�A�J�l�����c�ɑ���������̂́C�p���H�Ɛ��c���C���̗������C�r�����łȂ����Ă��āC�~�����Ɨ��������A�����Ă��邩�炾�Ɛ������Ă��܂��D���̂��Ƃ���l����ƁC�ŋߐ��c�n�тŃ~���}�A�J�l�̌������������̂́C�ޏꐮ�����ɂ���Ă��̂悤�Ȑ�����������������ƍl������ł��傤�D
�ʐ^�R�R�D�K�����������ł͂Ȃ����̂́C�~���}�A�J�l�́C��ł��C���̂悤�ɐ������܂����C����̂Ȃ��Ƃ�����D��ŎY������D
�@�~�����̃A�J�l���̒��ŁC��⑼�ƈ�������ɐ�������̂��C�q���A�J�l�ł��D�q���A�J�l�͎��n�̐����҂ł��D���U��ʂ��āC���n�Ƃ��̎��ӂ̎��тŐ������Ă���悤�ł��D�����Ȏ��n�����łȂ��C�r�̎��ӂɐ����������āC�������P��I�Ɏ��n��ԂɂȂ��Ă���C�Z�ݒ����Ă��邱�Ƃ�����܂��D�����Ȃ����n�͊J������Ă��܂����Ƃ������ł����C���R������X�ь����̒��Ɏ�荞�܂�Ă��鎼�n�������C�������������ł́C��������肵���̌Q���ێ��������̂Ǝv���܂��D
�ʐ^�R�S�D�q���A�J�l�͎��n���ɂ��������т��Đ������Ă���D���ߒr�ł��ݕӂ����n��ɂȂ��Ă���ΐ������Ă��邱�Ƃ�����D
�@�����ȊO�̃A�J�l���́C�Y���ꏊ�̊������قȂ���̂́C�ǂ��炩�Ƃ����Ε��n�ɂ��閾�邭�~�����ɑ����W�܂��Ă��܂�(�ʐ^�R�T)�D�ŋ�Y���������́C�Y�����ɐ��̂Ȃ��ꏊ�ɎY�����܂��D��ōl�@�����悤�ɁC����́C���̋x�����ƊW������̂�������܂���D�����Ă����̗c���͎Y���ꏊ�ɐڑ����Ă���~���ɐ������Ă��܂��D�������C�Ő��Y�������͂��̐���ɁC�œD�Y�������͂��̏ꏊ�ɐڑ����鐅��ɁC�c���͐������Ă��܂��D�����́C�Y���̊ώ@�ƁC���̐���ł̗c���̍̏W�o���ɂ���ė��t�����Ă���Ƃ����Ă悢�ł��傤�D�~������D�ނ̂́C�t���珉�Ăɂ����Đ������㏸���₷���C������������c���̔��炪�������邩�炾�Ƃ��l�����܂��D
�ʐ^�R�T�D�A�J�l���̐����������W�܂�Y������C�T�^�I�Ȑ����D
�@�ŋ�C�Ő��C�œD�̂��ׂĂ̎Y���l��������ԍ\���������āC���˂��炪���݂���D
�ʐ^�R�U�D�Ő��Y��������R�m�V���g���{(��)�ƁC���ۂőœD�Y������}�C�R�A�J�l(�E)�D�ʐ^�R�T�Ɠ����ꏊ�D
�ʐ^�R�V�D���ӎ��т̉��ŋx������m�V���g���{�̃I�X(��)�ƁC����ɐڑ����鑐�n�őŋ�Y������}�_���i�j���g���{(�E)�D�ʐ^�R�T�Ɠ����ꏊ�D
�@�ʐ^�R�T�̒r(���m�ɂ͂��̏ꏊ�̒r���Q)�ɂ́C���Ɍ��암�ɐ������Ă���A�J�l���̂قƂ�ǂ̎킪�����܂��D�������������������邭�J�����r���C���тɈ͂܂ꂽ�r���D�ރA�J�l���̎�����܂��D�i�j���g���{�C���X�A�J�l�������ł��D�����́C9���ɓ���������̏��������ɂ͓��ɁC�����Ƃ�����Ċݕӂ����n�����������Ɏ��т������킳���Ă���悤�ȏꏊ�őŋ�Y�������܂��D
�@�܂��C�L�g���{�́C�ʐ^�R�T�̂悤�Ȑr�ł͐��ۂœƓ��̑Ő��|�œD�Y��������̂́C�[���r�̒����ł��A���Ő��Y�����邱�Ƃ�����܂��D�L�g���{�͐����ꏊ�̐��ʕω��ɑΉ��ł���Y����p�������Ă���_�ŋ����[���g���{�ł��D
�ʐ^�R�W�D�i�j���g���{��X�A�J�l���D�ގY�����D�����Ƃ����ꂽ�r�݂̊Ɏ��т������킳���ĉA�ɂȂ��Ă���D
�ʐ^�R�X�D���Â����т̉��̗��㕔���őŋ�Y�����郊�X�A�J�l(��)�ƃi�j���g���{(�E)�D�ʐ^�R�W�̒r�D
�ʐ^�S�O�D�L�g���{�̎Y����p�D(��)���ʂ��Ⴂ�Ƃ��́C���ۂ̊ݕӂɓy�łł��������ȕǂɌ������đœD�Y�����Ă���D
�@(�E)���ʂ������ݕӂɓy�̕���������Ȃ��Ƃ����Ƃ��͒r�̒����őŐ��Y������D
�@�A�J�l���̃g���{�́C��ɏЉ���~���}�A�J�l�Ɍ��炸�C�c���n�тɑ����g���{�Ƃ�����ۂ�����܂��D�c���n�тɂ́C���ߒr�C�p���H�C�G�ؗтȂǁC�A�J�l���̐����j���ׂĂ��J�o�[����������ׂĂ�����Ă��邩��ł��D����ɐ��c���̂��̂��ꏊ�ɂ��Ă���A�J�l���̎�����܂��D���Ɍ��ł́C�i�c�A�J�l�������Ƃ��ڗ����C�A�L�A�J�l�C�~���}�A�J�l�Ȃǂ����c�ő��������܂����D�ߋ��`�ŏ������̂́C�A�L�A�J�l�͐����̂��������Ă��邱�ƁC�~���}�A�J�l�͏�ɏq�ׂ��悤�ɓc���n�т���قƂ�ǎp�������Ă��邱�ƁC�����ăi�c�A�J�l�́C���܂��ɐ��c�ŎY�����Ă���p������Ȃ�Ɍ���������̂́C�c��������Ă��邩�ǂ����͕�����Ȃ�����ł��D�Ƃ����̂��C���c�ł̃g���{�����ɁC��܂̉e�����w�E����Ă��邩��ł�(�Ⴆ�C�_�{�����C2009)�D
�@�ԂƂ�ڂ����{�l�ɂȂ��ݐ[�������̂́C�������������c�����҂��͂��߂Ƃ����A�J�l���̃g���{�������C�`���I�Ȉ�삪�s������ɓK�����Ă��āC���{�l�̐����̈�ɂǂ��Ɠ��荞����ł��傤�D
�ʐ^�S�P�D���Ɍ��ł̓A�L�A�J�l(���G2010.10.17.)��i�c�A�J�l(�E�G2009.9.26.)�Ȃǂ���Ȑ��c�����҂ł������D
�@���F�A�L�A�J�l�͈�肪�I��莼�n��ɂȂ������c�őœD�Y������C�E�F�i�c�A�J�l�͈��O�̑�����Ԃ̐��c�őŋ�Y������D
�@�ł́C�q�g�������n�߂�O�C�A�J�l���̃g���{�����͂ǂ����������Ő������Ă����̂ł��傤���D�����ĂȂ��C�`���I�Ȉ�삪�s������C������u���R�v�ɑ���������悤�ɂȂ����̂ł��傤���D�����͑z�����邵������܂��C�A�J�l���̐����j�̘g�g�݂⌻�݂̐����ꏊ�����Ƃɍl���Ă݂����Ǝv���܂��D
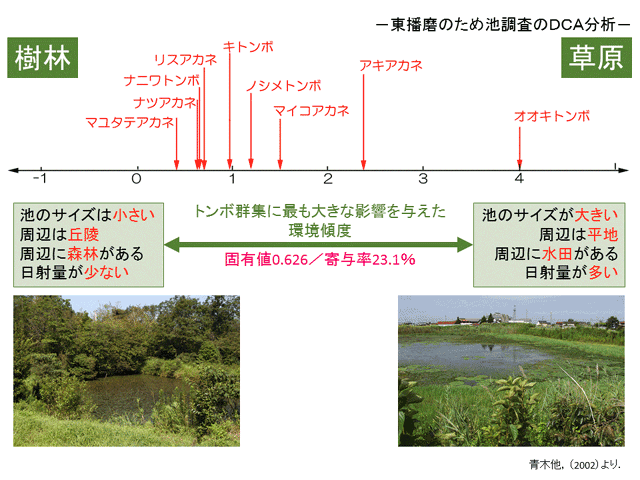
�@�A�J�l���̃g���{�̐����ꏊ�ƈꌾ�ł����Ă��C���łɌ��Ă����悤�ɁC���ꂼ��̎�ňقȂ�����D��ł��邱�Ƃ͊m���ł��D�����ŁC�A�J�l���Ɍ��炸�C���ߒr�̃g���{�Q�W�����肷����v���ׂ邱�Ƃ�ړI�Ƃ��āC2000�N����2001�N�ɂ����āC���̈قȂ镺�Ɍ��암�̂��ߒr35��I�肵�������܂����D�����́C�����̃g���{�Q�W�ƁC���ߒr�y�т��̎��ӂ̊��v�f���ʂ��C���̌��ʂ𑽕ϗʉ��(DCA)�ɂ����ĉ�͂��܂���(�������������C2006)�D���̌��ʂ����̐}�R�ł��D
�}�R�DDCA��ꎲ�̊��X�x�ƃA�J�l���e��̈ʒu�D���O�̂Ȃ����͔̂�������Ă��Ȃ��D
�}��4�̊��v�f���C�g���{�Q�W�̕ω��Ƒ��ւ�����Ǝ����ꂽ�D���Ȃ݂ɁC��͐����A���̑��l�������X�x�Ƃ��đI�ꂽ�D����̓g���{�̎Y���l���ƊW����ƍl������D
�������������̐������l�������F������������(2006)�}10a �y�ѐؑ�(2002)����D
�@���̌��ʂ�����Ǝ��̂悤�Ȃ��Ƃ�������܂��D�g���{�Q�W�������Ƃ��傫���ς�����v���́C�r�̃T�C�Y�C���n�����n���u�˂��C���ӂ����c���X�т��C���˗ʂ����������Ȃ����Ƃ������Ƃł��D�����4�̗v�f�͂��ꂼ��֘A���Ă���\��������܂��D�܂�C���c������ɂ��镽�n�̑傫�Ȃ��ߒr�͓��˗ʂ�����(�ȉ����n�r)�C�X�тň͂܂ꂽ�u�˒n�̏����Ȃ��ߒr�͓��˗ʂ����Ȃ�(�ȉ��u�˒r)�C�Ƃ������Ƃł��D
�@���̊��X�x�ɉ����Č�����A�J�l���̎���݂�ƁC�I�I�L�g���{�C�A�L�A�J�l�Ȃǂ����n�r�ɑ��������(���n���̎�)�C�}���^�e�A�J�l�C�i�c�A�J�l�C�i�j���g���{�C���X�A�J�l�Ȃǂ��u�˒r�ɑ���������(�u�ː��̎�)���Ƃ�������܂��D�����́C�g���{���悭�ώ@���Ă��錤���Ƃ̊��o�Ƃقڈ�v���Ă��܂��D�L�g���{�C�m�V���g���{�C�}�C�R�A�J�l�͂��̒��ԓI�Ȓr�Ƃ������ƂɂȂ邩������܂��C���̊ώ@����́C�m�V���g���{�ƃ}�C�R�A�J�l�͕��n�r�ɑ�������C�L�g���{�͂ǂ���ɂł��o������Ƃ�����ۂ������Ă��܂��D����Ɉ�t�������Ă����܂��ƁC���c�̐����҂Ƃ��ďЉ���i�c�A�J�l�́C�ǂ��炩�Ƃ����ΎR�Ԃ̐��c�ɑ����o�����܂��D���ߒr�Ō���ꂽ�i�c�A�J�l�͂���炪�x�����Ă���̂��Ǝv���܂��̂ŁC���̂悤�Ȍ��ʂ��o�Ă�����̂ƍl���Ă��܂��D
�@���̌��ʂł�������ڂ��ׂ��́C���n���̎�ł���I�I�L�g���{��A�L�A�J�l���C���n����̈ړ����������C�H�������r�̎��͂őO���B���̐������܂�������Ȃ��g���{�ł���C�Ƃ����_�ł��D�����āC�u�ː��̎�ł���}���^�e�A�J�l�C�i�j���g���{�C���X�A�J�l���H�������r���炻��قǑ傫���͈ړ����Ȃ��g���{�ł���Ƃ����_�ł��D���n�r�Ő������邽�߂ɂ́C�Ă��߂������߂̗������ꏊ�ֈړ�����͂��K�v�Ƃ���܂��D���������Ă����̃g���{�̈ړ����̍����́C���n�r�ł̐����ɓK�����Ă���Ƃ����܂��D����u�ː��̎�ɂ́C�Ă��߂����X�т��߂��ɂ���̂ŁC���̕K�v���Ȃ��Ƃ�����ł��傤�D
�@�Ƃ���ŁC�g���{�̗��z�~��}���ɐ�������c���ɂ��āCCorbet(1962)�͎��̂悤�Ȍ������q�ׂĂ��܂��D�܂��g���{�͔M�тŐi�������ܓx�n���֕��z���g�債�Ă������ƑO�Ă��܂��D���̏�ŁC���z�~�ɂ��ẮC�M�ђn���ɂ����Ċ��G�𗑂ŏ���Ƃ��������j�̘g�g�݂��C���ђn���ł̗��z�~�̑O�K��
*1�Ƃ��đ��݂��Ă����ƍl���Ă��܂��D�܂��Z���J�G�̊Ԃɐ�����悤�Ȉꎞ�I�Ȑ����܂�ɐ��炷��g���{�̗c���́C��ʂɉ��x�ɑ��锭�瑬�x�W���������C�������x�̂��Ƃŋ}���ɐ������鐫���������Ă���Ƃ��q�ׂĂ��܂��D�܂�C���G���~�ɁC�J�G���~�J�ɑΉ����āC�����j�̘g�g�݂��p����Ă��邢��Ƃ������Ƃł��D�A�J�l�������̘g�g�݂��p���ł���̂�������܂���D
�@�ł́C���{�Ŕ_�k���n�܂�O�̂悤����z�����Ă݂܂��傤�D���{�͐X�уo�C�I�[����L���鍑�ŁC���Ɍ��͏Ɨt���тɑ����܂��D�X�т͖L���Ȑ���ې����܂�����C�N����C������C�㗬��C������C������ƁC�͐�͏�ɖL���Ȑ����^��ł����Ƒz���ł��܂��D�܂��C��̐��������~�߂��Ăł����r��C�N���r�Ȃǂ̎~��������肵�����悾�����ł��傤�D�������������肵������ɂ́C�����炭�u�ː��̎킪�������Ă����Ƒz���ł��܂��D����ŁC���̐����j�̘g�g�݂������Ƃ��L���ɓ����C�܂�C��ԋ����̑���ɑ��ėL���ɓ����ꏊ�́C�͐�ɕt�����鎼�n�т�C�×����Ɍ`�������Ꮌ�n��ꎞ�I�Ȑ���̂悤�ȏ��ł͂Ȃ����Ǝv���܂��D�������������́C�Ă̊ԁC�͐�̐��ʂ�����������C���オ�����肵�āC�����Ȃ��Ȃ�\���������ƍl�����܂��D�ł�����C�c���Œ����ԕ�炳�˂Ȃ�Ȃ��g���{�͐������邱�Ƃ��ł��܂���D���ʂ�����C���������~�J�����ɋ}���ɐ������I���邱�Ƃ̂ł���A�J�l���̓V���Ƃ�����ł��傤�D
�ʐ^�S�Q�D��̗���̉��ϑw�ɔ��B�����_�n�͎R�̘[�܂ōL�����Ă���D
�@�q�g���_�k���n�߂܂��ƁC�×����̉��ϕ����Ꮌ�n�����c�ɕς���Ă����C�₪�ēc���n�тƂȂ��Ă��������Ƃ͑z���ɓ����܂���D�����Đl�������ƂƂ��ɁC�J���n�͂ǂ�ǂ�L����C�R�����ɂ܂Ŏ��������Ƃł��傤�D�����ɂ́C����ȑO���炠�����N���r�₻�̑��̒r���������āC�_�n�Ɏ�荞�܂�Ă�������������܂���D�R�����̊J���n�ɂ͐��c�������C���̂��߂̂��ߒr(�J�r)��p���H����������āC�{�������ɑ��݂��Ȃ������������l�H�I�ɂ����Ă��������Ƃł��傤�D�u�ː��̎�͂�������ēc���n�тɎ�荞�܂�Ă����C���n���̎�͊J���n�̍L����ƂƂ��ɓc���n�тɕ��z���L���Ă��������̂Ǝv���܂��D�����ăA�J�l���̃g���{�́u���R�v�ƌĂ��q�g�̐����̈�ɐ[�����荞��ł������̂��Ƒz���ł��܂��D
�@���R�̊��ł�������M���ׂ����Ƃ́C�q�g�̊����ɂ�靘���ł��D���t��������C�q�g�ɂ��A���J�ڂ̒�~�ł��D������C�Ԕ��C���ߒr�̐����C�p���H�̓D�グ�C�����ĉ���萅�c�ň����邽�߂̖��N�̈ێ���ƁC�Ȃǂ́C���ׂĊ����E�����̐A���J�ڂ��~�߂邱�ƂɂȂ�܂��D����ɂ���āC�l�X�̎Y�����C���n����n���̉߂��������E�ؘR����̂���сC���̈ێ��ɂ��c���̐���Ȃǂ̊�������I�ɒ���܂��D�×����͝������傫�ȏꏊ�ƍl�����C�����������J�ړr���̊��ɓK�����������𑗂��Ă���A�J�l���̃g���{�ɂƂ��āC�q�g�ɂ�靘�����t�Ɂu�J�ړr���Ƃ������肵���v����������Ă���Ƃ����܂��D
�@�ȏオ�C�A�J�l���̃g���{�����R�ɑ������R���ƍl�����܂��D
*1. �O�K���Ƃ́C������Ői�������K���I�`�����C�V�������ł��K�������������Ƃ������D
�g���{�̐��Ԋw�@���m�O���t�F�A�J�g���{
���Ɍ��̃A�J�l���̌���
�@�ȏ�A�J�l���̊e��ɂ��āC���̐��Ԃ��l���Ă��܂����D�Ō�ɁC���Ɍ��̃A�J�l���̌���ɂ��ĊT�ς��Ă��������Ǝv���܂��D������������Ɍ��ɐ������Ă���15��̂����C��Ŋ뜜��Ƃ��ē��{�̃��b�h���X�g(2007)�ɍڂ��Ă�����̂�3�킢�܂��D��Ŋ뜜�T�ނ̃}�_���i�j���g���{�ƃI�I�L�g���{�C��Ŋ뜜�U�ނ̃i�j���g���{�ł��D�����͕��Ɍ��̃��b�h���X�g(2012)�ł��C���ꂼ��`�C�a�C�b�Ƀ����N����Ă��āC����Ƀ~���}�A�J�l���b�C�A�L�A�J�l�ƃq���A�J�l���v���ڎ�Ɏw�肳��Ă��܂��D
�@���̒��Ń}�_���i�j���g���{�������Ƃ���ł̉\����������ł��傤�D2018�N���݊m���Ȑ����n��1�J���ŁC�����ł��C�̐����������Ă��܂��D����2016�N�ɃI�X�̌̂��m�F���Ă��܂��D2017�N�͊ώ@�ɍs�����Ƃ��ł��܂���ł����D�}�_���i�j���g���{�ɂ��ďڂ�����
������̃y�[�W���������������D
�ʐ^�S�R�D���Ɍ��̃}�_���i�j���g���{�̐����n�ɂ���������Ƃ��ŋ߂̋L�^(2018�N6������)�D
�@���F���Ɍ��̂��Ă̑�Y�n�ɂ�����ŋ߂̋L�^�C2010.10.10�C�E�F���Ɍ��̍Ō�Ǝv����Y�n�̍ŋ߂̋L�^�C2016.10.10.�D
�@���āC�S���I�ɂ͂��Ȃ茸�����Ă���Ƃ����Ă���I�I�L�g���{�́C���Ɍ��ł͈��肵�Ċe�n�Ŗ��N�ώ@���邱�Ƃ��ł��܂��D���Ɍ��ɑ������R�́C���ߒr�̐����S���ꑽ���C����炪���Ɍ��암�ɏW�����Ă��邱�ƁC�ŋߓ��ɏH�ɂ��ߒr�̐������悭�s����悤�ɂȂ������ƂȂǂ��l�����܂��D�I�I�L�g���{�͐��ʂ̉����������ߒr�̐��ۂɍL����D�y�̕����ɑœD�Y�����܂��D�H�̐����ɂ���Ă��̂悤�ȎY�����������̂��ߒr�Ɍ��o���܂����D�܂��H�������r����傫���ړ����U���Đ������C�Y���V�[�Y���ɂȂ�ƍĂшړ����ĐV���ȎY������L���邽�ߒr��T���K�������邱�Ƃ���C���ߒr�̑������C���̔N�X�ŕω�����Y���ꏊ�������₷�����Ă���ƍl�����܂��D�����Ă���͐����ł����C�����Ƃ����������C�×����Ői�����Ă����Ǝv����I�I�L�g���{�ɗL���ɓ����Ă���̂�������܂���D�I�I�L�g���{�ɂ��ẮC�������̒r�ɏZ�ݒ����Ƃ����̂ł͂Ȃ��C�L��������Ԃ̒��ɂ�������̂��ߒr�������āC���̒�����K�ȎY�����̂���r��I��Ő������Ă���C�Ƃ������o�ő�����Ƃ悢�ł��傤�D
�@������������S�z�Ȃ��Ƃ�����܂��D����͂��ߒr�ւ̑��z�����d�{�݂̐ݒu�ł��D��������̂悢���ߒr�C�܂��ɂ��ꂪ�I�I�L�g���{�̍D�ޒr�ł����C�����ɑ��z�����d�{�݂����������n�߂Ă��܂��D���ꂪ�I�I�L�g���{�ɂǂ̂悤�ȉe����^���邩�́C����ώ@�𑱂��Č�����Ă����K�v������ł��傤�D

�ʐ^�S�S�D�������Ƃ��ꂽ���ɓK�����Ă���I�I�L�g���{�D���̋�������ɑ��ėL���ł��邩������Ȃ��D�Y���ʐ^�͓����r�D
�ʐ^�S�T�D���n�̐�������̂悢�r�ɐݒu����Ă��郁�K�\�[���[���d�{�݁D
�@�i�j���g���{�ɂ��Ă��C�I�I�L�g���{���l�C���ߒr�̑����ƁC�H�̐������悭�����Ă���Ǝv���܂��D�����C�I�I�L�g���{���ǂ��炩�Ƃ����ΊJ�������n�r(������M�r)�ɑ����W�܂�̂ɑ��C�i�j���g���{�͎��тɈ͂܂ꂽ�u�˒r(������J�r)�ɑ����Ƃ����Ⴂ�͂���܂��D������ɂ��Ă��C�ڂ�����
������̃y�[�W���������������D
�@���Ɍ��̃~���}�A�J�l���������������g���{�ł��D����ɂ��Ă͂��łɍl�@�����悤�ɁC�~�����Ɨ��������A�������悤�ȏꏊ�ŗc������Ƃ������Ƃ���C���̂悤�Ȋ��̌����C���ɓc���n�т̕ޏꐮ���ɂ����̂��C���������ƍl�����܂��D���݃~���}�A�J�l��������ꏊ�́C���̑�������ł��D��̈ꕔ�ɗ���ɂȂ��鏬�K�͂Ȏ~���I�������o���Ă���悤�ȏꏊ������C���ꂪ�����N���ɂ킽���Ĉ��肵�Ă���悤�ȂƂ���ɐ����c���Ă���悤�Ɍ����܂��D�����������ꏊ�͂ƂĂ����Ȃ����C���K�͂ł��傤����C�̐��͂����������ꏊ�ł����ɏ��Ȃ��ƍl�����܂��D����1�C2�����ۂ�Ǝ~�܂��Ă���Ƃ������ώ@�Ⴊ�����Ȃ��Ă��܂��D
�ʐ^�S�U�D���F�ۂ��1�������R���t�߂Ɏ~�܂��Ă���~���}�A�J�l�D2010.8.22.�C�E�F���n�̎M�r�ɐ�������}�C�R�A�J�l�D2006.9.17.
�@���b�h���X�g�Ɍf�ڂ���Ă��Ȃ���ŁC����Ԍ����X�����������Ǝv����̂��C�}�C�R�A�J�l�ł�(cf. �C2014)�D�������ł����l�̕��Ȃ���Ă��܂�(�k�R���C2018)�D���̊ώ@�ł́C�ߋ��ɋL�^�̂����������n��K��Ă��C���݂قƂ�ǂ��̎p�������Ă��܂��D��̂��ߒr�������s��ꂽ2001�N����Ɋe�n�Ō���ꂽ�}�C�R�A�J�l�́C�}�R�̕��͌��ʂ�����C�ǂ��炩�Ƃ����Ε��n�r�ɋ߂����ɑ����Ƃ������ʂ��o�Ă��܂��D���ہC��������̂悢���n�̎M�r�ɍL����q���K�}�̌Q���̒��ŁC���n���ォ�琬�n����܂ʼn߂����Ă���̂��ώ@�������Ƃ�����܂���(�ʐ^�S�U�E)�D�����}�C�R�A�J�l�̌����̌����́C���̂Ƃ���s���ł��D����ł������Ă����Ȃ�C�ŋߕ��n�r�ɒ蒅���̂���g���{�Q�W���n��ɂȂ��Ă������ۂ��C�e�n�̊ώ@��ʂ��Ď����Ă���C�}�C�R�A�J�l�����̈�Ō������Ă���̂ł͂Ȃ����Ɗ����Ă��܂��D
�@�����ȊO�ł́C�A�L�A�J�l�̌������C���ɕ��Ɍ��암�ŔF�߂��܂��D���Ɍ���_�ˎs�ł̏�
������̃y�[�W�ɕ��܂����̂ł������������D����ɂ��ẮC�悭�����Ă���悤�ɁC��܂̉e�����傫���Ǝv���܂��D���ɁC���_��E���_��̒n��ł́C�̂̂悤�ɔ��̑����̃A�L�A�J�l�����݂����c�𗘗p���������Ă��܂��D
�@�A�L�A�J�l�Ɣ���Ⴗ��悤�ɁC�ŋ߂悭�p������悤�ɂȂ����̂̓^�C���N�A�J�l�ł��D�^�C���N�A�J�l�̔ɐB�������̌̂́C�ȑO�́u�������ɂ͔�������Ȃ��v�ƕ���Ă��܂���(���ԁC1969)�D�������C���̒���́C���̊ώ@�ɂ���Ă��C�C�����K�v�ɂȂ��Ă���Ƃ�����ł��傤�D�ŋ߂͊C�݂���20km�ȏ�����ꂽ�������̒r���ŁC�H�ɂ�������̌̂��W�����ĎY���������s���Ă��܂��D�������I�I�L�g���{�̎Y���Əd�Ȃ�C�I�I�L�g���{�Ƃَ̈�ԘA����������قǂł��D�^�C���N�A�J�l�͑Ő��Y�����s���C�w�Z�̃v�[���ł��c�����悭������܂����C�������̊ώ@�ł́C�ނ��됅�ۂŁC�œD�Y���ɋ߂��Y�����s���Ă���̂����܂��D�؋��͂܂���������܂��C�A�L�A�J�l�̌������C��������^�C���N�A�J�l�̓������ւ̐i�o���\�ɂ����ƍl���邱�Ƃ͂ł��Ȃ��ł��傤���D

�ʐ^�S�V�D���F�C�݂���20km�ȏ�������̒r�ɂ�������W�܂��ĎY������^�C���N�A�J�l�����D���ۂőœD�Y�����Ă���̂�������D
�@�E�F�I�I�L�g���{�̃��X�ƈَ�ԘA�����Ă���^�C���N�A�J�l�̃I�X�D�I�I�L�g���{�Ɠ����r�ŎY�����Ă���̂��D�Ƃ���2014.10.26.
�@�ŋ߂̕��n�r�ł̃A�J�l���̊ώ@�ł́C�I�I�L�g���{��^�C���N�A�J�l���ڗ��̂ł����C��������ړ����̍�����ł��D����ŕ��n�r�Ƃ��̎��ӂňꐶ�𑗂�悤�ȃ}�C�R�A�J�l�C�A�J�l���ł͂���܂��R�t�L�g���{�Ȃǂ��������Ă��鎖��������܂��D����͒r�₻�̎��ӂ̊��������������Ă��܂��D�L���ړ����邱�ƂŎY���ꏊ��I���ł���킾�����C�r�������̃��X�N�U�����邱�ƂɂȂ��Ă���̂�������܂���D
�@�ȏ�q�ׂ��ȊO�̃A�J�l���́C�S�̂Ƃ��Č����X������������̂́C�܂���@�I�ɂ܂ł͎����Ă��Ȃ��Ƃ�����ł��傤�D
�Q�l����
����k��C1988�D�d���̍����D�_�ːV�������o�ŃZ���^�[�C�_��.
�ؓT�i�C2014�D���Ɍ��ɂ����āC�g���{�����n�Ƃ��Ď{�݁E���������ʂ����Ă�������DAeschna 50: 33-38.
�ؓT�i�C�p��N�Y�C�O���O�@�C�c���N�v�C����r���C����b�C�����T�q�D2002�D���ߒr���琶�����l���̕ۑS���l���� (4)�g���{�Q�W�����߂�v���D���{�����w���67����u���v�|�W:193.
���P��C2010�D���Ɍ��̃g���{���z�ژ^�i���P��̋L�^1958-2007�j�DSympetrum Hyogo, 121-113.
Hiroshi Ando, 1957. A comparative study in the development ommatidia in Odonata. Sci. Rep. Tokyo Kyoiku Diag.(B) 8:174-216.
Hiroshi Ando, 1962. The comparative embryology of Odonata with special reference to a relic dragonfly Epiophlebia superstes Selys. Jap. Soc. Prom. Sci., Tokyo.
��c�N�s�C1990�D�g���{�ނ�"autumn species"�̐����j�D�̌Q���Ԋw���� 46:62-67�D
��c�N�s�C1993�D�R�֏��A�L�A�J�l�C���Ȃ��A�L�A�J�l�@�A�L�A�J�l�̐����j�ɂ����鏔���|�P�D�C���Z�N�^���E�� 30�F292-299�D
��c�N�s�C1996�D�A�J�g���{�ނ̑��l�ȔɐB�s�����ǂ��������邩�D�����Ǝ��R 31(8):2-8.
���ԖC1967�D�^�C���N�A�J�l�̎Y������t���܂ŁDGracile 5:7-10.
���ԖC1969�D���t�߂ɉ�����^�C���N�A�J�l�̐��ԁDTombo 12(1/4):17-23.
�k�R��E�ѐ����E��l�ˎ��C2018�D�������ɐ�������g���{�ڂ̖ژ^�Ɛ��Ԏʐ^�W�D�z�V�U�L�O���[�����c�������ʍ��D��22���D116pp. ���v���c�@�l�z�V�U�L�O���[�����c�C�z�V�U�L�쐶�����������D
�������������C2006�D�������l���̌����@�\�̉𖾂ƕۑS�v���W�F�N�g(�I����)�@����13�`17�N�x�D���������������ʌ�����, SR-72-2006�C 81pp.
P. S. Corbet, 1962. A biology of dragonflies. H. F. & G. Witherby ltd., London.
Matsura, T. K.Komatsu K.Komatsu K.Nomura & M.Oh'Oto, 1995. Life history of Sympetrum striolatum imitoides Bartenef at an outdoor swimming pool in an urban area (Anisoptera Libellulidae). Odonatologica 24(3):291-300.
�ߓ��ˎq�C1994�D����̃g���{�̗����E�c�����̋L�^�DAeschna 29:15-20.
�ߓ��ˎq�C1995�D����̃g���{�̗����E�c�����̋L�^�D�lj��P�DAeschna 31:7-10.
�V������E�ߓ��ˎq�C1989�D�A�J�g���{�̗����y�ї��̑傫���ɂ��āDGracile 42:1-5.
�_�{�����C��c�N�s�C�܉ӌ���C������C���Ǐr���C2009�D�t�B�v���j���ƃC�~�_�N���v���h�𐬕��Ƃ����c���{�p�E���܂��A�L�A�J�l�̗c���ƉH���ɋy�ڂ��e���D�_�Ɣ_���H�w��_���W, 259:35-41.
�������r�E���O�s�C1990�D��g�W���j�A�V��178�C�g���{�����ւ悤�����D��g���X�C����.
�n������Y�C1953�D�V�����k���n������x(��O��) �A�L�A�J�l�̐��ԑ��̑��D�z���������D���X�� 7(4)�F128-139�D
�l�c�N�E��㐴�C1985�D���{�Y�g���{��}�ӁD�u�k�ЁC�����D
Wataru Higashikawa, Mayumi Yoshimura, Tsuyoshi Yagi & Kaoru Maeto, 2016. Microhabitat use by larvae of the endangered dragonfly Sympetrum pedemontanum elatum (Selys) in Japan. J Insect Conserv 20:407-416.
Fraser, F. C., 1957. A Reclassification of the Order Odonata. Royal Zoological Society of New South Wales.
Sinzo Masaki, 1980. Summer Diapause. Ann. Rev. Entomol. 25:1-25.
�{��r�s�C2018�D�i�j���g���{��7���̎Y����DGracile 78:17-18.
�R�{�N���E�V������E�{��r�s�E���Y�M���C2009�D�ߋE�̃g���{�}�ӁD�������ЁC�����D
�n�ӗf�q�C1998�D�A�J�g���{�̗������Ǝ��c���ɂ��āD�����Ǝ��R 31(8):13-17.