私は,神戸市内のトンボの調査を始めたころ,5月下旬に,キイロサナエが同時にたくさん羽化している姿を目にしました.この同調的羽化はしばらく続きましたが,2週間もするとパタッと羽化は見られなくなりました.調べてみると,キイロサナエの幼虫期間は2年から4年である(関西トンボ談話会,1984)と書かれていました.こんなに長い間幼虫で過ごしていて,どうして,5月下旬にだけ,しかもこのような短期間だけに,羽化が制限されるのだろう,という強い疑問を持ちました.しかしそのことについて解説している書籍や文献は見つかりませんでした.例えば,多くの解説書や図鑑には,「フタスジサナエは4月下旬に羽化が始まり,その後6月まで成虫が見られる」といったように,各トンボの出現季節は記されています.しかし,なぜその時期にだけ成虫が出現するのかといったしくみについては記述されていません.
トンボの生態学
7.生活史の季節的制御 目次
- トンボの生活史の分類
Ⅰ.成虫出現季節と生活史の分類
Ⅱ.成虫越冬種の生活史
Ⅲ.春季種の生活史
Ⅳ.夏季種の生活史
Ⅴ.秋季種の生活史 - トンボの休眠
Ⅰ.広義の休眠,休眠,休止,そして休眠発育
Ⅱ.卵の休眠
Ⅲ.幼虫の休眠
Ⅳ.成虫の休眠
トンボの生態学 7.生活史の季節的制御
トンボの生活史の分類

写真1.キイロサナエの同調的羽化,2018.5.26.羽化している個体のそばに羽化を終えて残された羽化殻がついている.
このしくみ,つまり特定のステージ(例えば成虫)が特定の季節に配置されるように調整されることは,一般に「季節的制御 (seasonal regulation)」と呼ばれています.
トンボの季節的制御のしくみについては,海外では,20世紀中頃から色々と研究が行われ,たくさんの論文が出ています.しかし日本では,ことトンボに関する限り,このような研究はわずかですし,論文も非常に数が少ないというのが現状です.日本にははっきりとした四季があり,日本人にも独特の季節感が存在すると思うのですが,この分野に興味を持ったトンボ研究者がほとんどいなかったというのは残念な限りです.他の昆虫分類群では,例えば正木進三博士のコオロギの季節的制御の研究は非常に有名で,一般向け解説書まで出ています(例えば,正木,1974).
この章では,主に日本のトンボに焦点を当てて,季節的制御について解説してみようと考えています.まずはトンボの成虫出現季節による生活史分類を行って,どのような生活史を持っているトンボがいるかというところから始めてみましょう.
Ⅰ.成虫出現季節と生活史の分類
たくさんのトンボを観察しようとすればすぐに気がつくことですが,種によって成虫の出現する季節が異なっています.春の一時期にしか成虫が見られない種,初夏から夏の暑い時期に多く見られる種,秋に目立ち始め繁殖活動をする種,ほぼ暖かい季節の間はずっと見られる種など,さまざまです.
成虫の出現季節からみると,日本のトンボの生活史は,大きく4つに分類できると考えられます.すなわち,夏に羽化しそのまま成虫で越冬して春に繁殖活動を行う種,春を中心とした一時期にだけ成虫が出現する種,種によって期間は異なるものの夏を中心に成虫が出現する種,羽化が初夏に行われ秋を中心に繁殖活動をする種です.それぞれ成虫越冬種(overwintering as adults;e.g., Norling & Sahlén,1997),春季種(spring species;Corbet,1962),夏季種(summer species;Corbet,1962),秋季種(autumn species;上田,1990)と呼ばれています.この4つの生活史のうち,夏季種の成虫出現期間がもっともバラエティに富んでいて,兵庫県でいえば,4月下旬から11月まで成虫が見られるシオカラトンボのような種から,6月下旬に姿を見せはじめ9月上旬には姿が見られなくなるネアカヨシヤンマのような種までを含みます.
これらの成虫の出現季節は,実は冬をどのステージで越すかという,トンボの越冬状態と非常に関連が深くなっています (例えば Corbet, 1962;Norling & Sahlén,1997).
まず成虫で越冬する種は,その名の通り,成虫が越冬ステージです.春季種と夏季種は幼虫で越冬します.秋季種は卵で越冬するのが基本で,個体群の一部が若齢幼虫で越冬している場合があります.春季種と夏季種の違いは,前者は翌年羽化する個体のほとんどが終齢で越冬するのに対し,後者は翌年羽化する個体が終齢を含むさまざまの齢で越冬しているという違いがあります.その結果,幼虫の成長が進んでいてかつそろっている春季種は春の一時期にだけ成虫が出現し,幼虫の成長にばらつきがある夏季種は出現期間が一般に長くそして遅くなる(夏になる)ということになります.
トンボの季節的制御のしくみについては,海外では,20世紀中頃から色々と研究が行われ,たくさんの論文が出ています.しかし日本では,ことトンボに関する限り,このような研究はわずかですし,論文も非常に数が少ないというのが現状です.日本にははっきりとした四季があり,日本人にも独特の季節感が存在すると思うのですが,この分野に興味を持ったトンボ研究者がほとんどいなかったというのは残念な限りです.他の昆虫分類群では,例えば正木進三博士のコオロギの季節的制御の研究は非常に有名で,一般向け解説書まで出ています(例えば,正木,1974).
この章では,主に日本のトンボに焦点を当てて,季節的制御について解説してみようと考えています.まずはトンボの成虫出現季節による生活史分類を行って,どのような生活史を持っているトンボがいるかというところから始めてみましょう.
Ⅰ.成虫出現季節と生活史の分類
たくさんのトンボを観察しようとすればすぐに気がつくことですが,種によって成虫の出現する季節が異なっています.春の一時期にしか成虫が見られない種,初夏から夏の暑い時期に多く見られる種,秋に目立ち始め繁殖活動をする種,ほぼ暖かい季節の間はずっと見られる種など,さまざまです.
成虫の出現季節からみると,日本のトンボの生活史は,大きく4つに分類できると考えられます.すなわち,夏に羽化しそのまま成虫で越冬して春に繁殖活動を行う種,春を中心とした一時期にだけ成虫が出現する種,種によって期間は異なるものの夏を中心に成虫が出現する種,羽化が初夏に行われ秋を中心に繁殖活動をする種です.それぞれ成虫越冬種(overwintering as adults;e.g., Norling & Sahlén,1997),春季種(spring species;Corbet,1962),夏季種(summer species;Corbet,1962),秋季種(autumn species;上田,1990)と呼ばれています.この4つの生活史のうち,夏季種の成虫出現期間がもっともバラエティに富んでいて,兵庫県でいえば,4月下旬から11月まで成虫が見られるシオカラトンボのような種から,6月下旬に姿を見せはじめ9月上旬には姿が見られなくなるネアカヨシヤンマのような種までを含みます.
これらの成虫の出現季節は,実は冬をどのステージで越すかという,トンボの越冬状態と非常に関連が深くなっています (例えば Corbet, 1962;Norling & Sahlén,1997).
まず成虫で越冬する種は,その名の通り,成虫が越冬ステージです.春季種と夏季種は幼虫で越冬します.秋季種は卵で越冬するのが基本で,個体群の一部が若齢幼虫で越冬している場合があります.春季種と夏季種の違いは,前者は翌年羽化する個体のほとんどが終齢で越冬するのに対し,後者は翌年羽化する個体が終齢を含むさまざまの齢で越冬しているという違いがあります.その結果,幼虫の成長が進んでいてかつそろっている春季種は春の一時期にだけ成虫が出現し,幼虫の成長にばらつきがある夏季種は出現期間が一般に長くそして遅くなる(夏になる)ということになります.

写真2.成虫越冬種であるホソミオツネントンボ(左)と春季種であるフタスジサナエ(右).帯グラフの赤色部分が成虫出現期.

写真3.夏季種であるヤブヤンマ(左)と秋季種であるオオキトンボ(右).帯グラフの赤色部分が成虫出現期.
Ⅱ.成虫越冬種の生活史
日本には,成虫で越冬する種は3種知られています.アオイトトンボ科のオツネントンボ Sympecma paedisca,ホソミオツネントンボ Indolestes peregrinus,そしてイトトンボ科のホソミイトトンボ Aciagrion migratum です.
オツネントンボ(写真4左)は,兵庫県南部では,3月下旬には池に現れ,4月から5月中旬にかけて繁殖活動が行われます.卵と幼虫のステージはあわせて3ヶ月ほどで完了し,早いものでは6月下旬には次の世代の成虫が現れます.この成虫はその年のうちには繁殖活動を行わず,翌年の春を待って繁殖活動を開始します.
羽化後の夏,そして秋・冬の間,オツネントンボは繁殖活動を行いません.この繁殖活動の遅延を「生殖休眠 (reproductive diapause)」と呼んでいます(同じオツネントンボ属の Sympecma fusca について,Norling & Sahlén,1997).この生殖休眠が生理的にどのようなしくみで引き起こされているかについては知られていません.また生殖休眠がどのような合図で消去されるのかについても, 成虫越冬種については知見がありません.
ホソミオツネントンボ(写真2左)も,春の成虫の出現期間がやや長く,次の世代の出現が少し遅いことを除けば,オツネントンボとほぼ同じ生活史を営んでいます.

写真4.成虫越冬種.オツネントンボの産卵(左;2011.4.5.),ホソミイトトンボの産卵(右;2019.5.3.)
ホソミイトトンボ(写真4右)は,上記の2種とは違って,6月下旬頃に羽化した次世代の成虫は生殖休眠の状態にならず,そのまま夏に繁殖活動を行います.そしてこの次の,越冬する世代の成虫は,9月に入ってから羽化をしてきます.こちらは秋には繁殖活動を行わず,次の春まで持ち越されます.
Ⅲ.春季種の生活史
兵庫県南部の春季種は多くのサナエトンボ科に見られ,それ以外では,カワトンボ科のニホンカワトンボ Mnais costalis とアサヒナカワトンボ Mnais pruinosa,ヤンマ科のクロスジギンヤンマ Anax n. nigrofasciatus とサラサヤンマ Sarasaeschna pryeri,ムカシトンボ科のムカシトンボ Epiophlebia superstes,ムカシヤンマ科のムカシヤンマ Tanypteryx pryeri,エゾトンボ科のトラフトンボ Epitheca marginata,トンボ科のシオヤトンボ Orthetrum japonicum,ヨツボシトンボ Libellula quadrimaculata asahinai,ベッコウトンボ Libellula angelina,等々が該当します.
海外では,より北方に分布するアカメイトトンボ属,カラカネトンボ属,カオジロトンボ属等の種も春季種に含まれます(Norling & Sahlén, 1997).他にもあるかも知れませんが,個体群の大部分が終齢幼虫で羽化前の冬を越す,というのが春季種の正しい定義ですから(Corbet, 1962),幼虫の成長状態をきちんと把握しなければ確実なことはいえません.なお,Norling & Sahlén (1997) は,spring species という語の代わりに,early breeders (早期に繁殖活動をするもの)という語をあてています.
春季種には,幼虫期間が2年以上にわたる,多年一化*1の生活史を持つものが多いのが特徴です.サナエトンボ科に多く,ムカシトンボやムカシヤンマも多年一化です.北方では,カオジロトンボ原名亜種 Leucorrhinia dubia (Norling, 1976),カラカネトンボ(Ubukata, 1980)なども多年一化です.当然のことながら,多年一化の場合越冬幼虫の齢期は広い範囲に渡りますが,翌年羽化するのは,その中で終齢幼虫の個体たちだけということになります.多年一化以外のトンボは,すべて一年一化です.一年に二化以上するものは,spring species や eraly breeders の意味から考えて,あり得ません.
また,化性*2に関係なく,春季種の卵期間はだいたい40日以下です(関西トンボ談話会,1984).Ando (1962) は,トンボ21種の胚発生の研究から卵期間が40日以下のものを非休眠性種 (non-diapause species) の卵としていますし,Corbet (1999) はその後の知見を加えて60日以下のものを直接発生卵 (direct development eggs:停止することなく発生する卵の意味)としていますので,春に産下されたこれら春季種の卵はそのまま停滞することなく発生を続け,孵化に至ると考えてよいでしょう.
春季種は,翌春に羽化する個体たちが,終齢幼虫という成長のそろった状態で羽化前の冬を越すことから,羽化の同調性が非常に高くなっているのが特徴です.ある生息地の一年の羽化が,わずか3週間前後で終わってしまうこともあります(例えばキイロサナエ;Aoki,1999;右図).羽化の同調性を表す指標にEM50という数値があります.年間羽化数の50%が羽化 (EMergence) するのにかかる日数で表現されます.キイロサナエの場合,性差や年変動はあるものの,EM50の値は4日程度になります(Aoki, 1999).つまり一年に羽化してくる成虫の半数がわずか4日間で羽化してしまうのです.ものすごい同調性の高さです.ちなみに幼虫期間は3または4年という長期間です(Aoki, 1999).ここには,何か羽化の同調性を高めるしくみの存在があると考えざるを得ません.
Ⅲ.春季種の生活史
兵庫県南部の春季種は多くのサナエトンボ科に見られ,それ以外では,カワトンボ科のニホンカワトンボ Mnais costalis とアサヒナカワトンボ Mnais pruinosa,ヤンマ科のクロスジギンヤンマ Anax n. nigrofasciatus とサラサヤンマ Sarasaeschna pryeri,ムカシトンボ科のムカシトンボ Epiophlebia superstes,ムカシヤンマ科のムカシヤンマ Tanypteryx pryeri,エゾトンボ科のトラフトンボ Epitheca marginata,トンボ科のシオヤトンボ Orthetrum japonicum,ヨツボシトンボ Libellula quadrimaculata asahinai,ベッコウトンボ Libellula angelina,等々が該当します.
海外では,より北方に分布するアカメイトトンボ属,カラカネトンボ属,カオジロトンボ属等の種も春季種に含まれます(Norling & Sahlén, 1997).他にもあるかも知れませんが,個体群の大部分が終齢幼虫で羽化前の冬を越す,というのが春季種の正しい定義ですから(Corbet, 1962),幼虫の成長状態をきちんと把握しなければ確実なことはいえません.なお,Norling & Sahlén (1997) は,spring species という語の代わりに,early breeders (早期に繁殖活動をするもの)という語をあてています.
春季種には,幼虫期間が2年以上にわたる,多年一化*1の生活史を持つものが多いのが特徴です.サナエトンボ科に多く,ムカシトンボやムカシヤンマも多年一化です.北方では,カオジロトンボ原名亜種 Leucorrhinia dubia (Norling, 1976),カラカネトンボ(Ubukata, 1980)なども多年一化です.当然のことながら,多年一化の場合越冬幼虫の齢期は広い範囲に渡りますが,翌年羽化するのは,その中で終齢幼虫の個体たちだけということになります.多年一化以外のトンボは,すべて一年一化です.一年に二化以上するものは,spring species や eraly breeders の意味から考えて,あり得ません.
また,化性*2に関係なく,春季種の卵期間はだいたい40日以下です(関西トンボ談話会,1984).Ando (1962) は,トンボ21種の胚発生の研究から卵期間が40日以下のものを非休眠性種 (non-diapause species) の卵としていますし,Corbet (1999) はその後の知見を加えて60日以下のものを直接発生卵 (direct development eggs:停止することなく発生する卵の意味)としていますので,春に産下されたこれら春季種の卵はそのまま停滞することなく発生を続け,孵化に至ると考えてよいでしょう.
春季種は,翌春に羽化する個体たちが,終齢幼虫という成長のそろった状態で羽化前の冬を越すことから,羽化の同調性が非常に高くなっているのが特徴です.ある生息地の一年の羽化が,わずか3週間前後で終わってしまうこともあります(例えばキイロサナエ;Aoki,1999;右図).羽化の同調性を表す指標にEM50という数値があります.年間羽化数の50%が羽化 (EMergence) するのにかかる日数で表現されます.キイロサナエの場合,性差や年変動はあるものの,EM50の値は4日程度になります(Aoki, 1999).つまり一年に羽化してくる成虫の半数がわずか4日間で羽化してしまうのです.ものすごい同調性の高さです.ちなみに幼虫期間は3または4年という長期間です(Aoki, 1999).ここには,何か羽化の同調性を高めるしくみの存在があると考えざるを得ません.
*1.多年一化 partvoltine.産卵から羽化まで2年以上かかる生活史をいう.
*2.化性 voltinism.昆虫が一年に繰り返す世代の数.
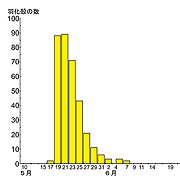
図1.キイロサナエの羽化曲線(Aoki, 1999)

写真5.春,同調的に羽化するフタスジサナエ,2018.4.20.春季種は羽化の同調性が非常に高い.
Ⅳ.夏季種の生活史
夏季種は翌年羽化する個体たちがさまざまの齢期で幼虫越冬する種と定義されます.多くのトンボがこの範疇に入ると考えられますが,それらを列記するより,春季種,秋季種,成虫越冬種を除くすべてのトンボ,と考える方が分かりやすいでしょう.
夏季種の化性は様々です.一年二化(あるいはそれ以上),一年一化,多年一化の種が存在します.以下,兵庫県地域での化性について,筆者の幼虫採集経験を中心に,尾園ら(2012)を参考にしながら概観してみましょう.
一年二化あるいは多化している種には,アオモンイトトンボ Ischnura senegalensis やオオイトトンボ Paracercion sieboldii を初めとしたイトトンボ科の多くの種,ヤンマ科のギンヤンマ Anax parthenope julius,シオカラトンボ Orthetrum albistylum speciosum やショウジョウトンボ Crocothemis servilia mariannae などのトンボ科の数種などが該当します.時に二化目の個体のサイズが一化目より小さくなることがあります(写真6右).また一年に二化以上する種は,春比較的早い時期に一化目の成虫が現れ,没姿時期もかなり遅くなる傾向があります.これらはより低い温度に適応している種たちなのかも知れません.あるいはまた,同じ温度に対して成長速度が高い種なのかも知れません.

写真6.一化目と二化目のサイズの違い.左はアオモンイトトンボのメス.一化目のサイズの大きい個体が二化目の小さい個体に食いついている.
右はコフキトンボの羽化殻.左側は6月15日採集の一化目,右側は8月20日採集の二化目.明らかに二化目の方が小さい.
右はコフキトンボの羽化殻.左側は6月15日採集の一化目,右側は8月20日採集の二化目.明らかに二化目の方が小さい.
一年一化の夏季種は,初夏や夏を中心に成虫が出現する種に多いようです.例えば,カワトンボ科ではアオハダトンボ Calopteryx japonica やハグロトンボ Atrocalopteryx atrata,モノサシトンボ科ではグンバイトンボ Platycnemis foliacea sasakii,イトトンボ科ではベニイトトンボ Ceriagrion nipponicum やヒヌマイトトンボ Mortonagrion hirosei,ヤンマ科ではヤブヤンマ Polycanthagyna melanictera やマルタンヤンマ Anaciaeschna martini,トンボ科ではコシアキトンボ Pseudothemis zonata などが該当します.
多年一化の種としては,オニヤンマ科のオニヤンマ Anotogaster sieboldii,サナエトンボ科のウチワヤンマ Sinictinogomphus clavatus clavatus,ヤンマ科のコシボソヤンマ Boyeria maclachlani やミルンヤンマ Planaeschna milnei milnei,エゾトンボ科のエゾトンボ Somatochlora viridiaenea やハネビロエゾトンボ Somatochlora clavata,ヤマトンボ科のオオヤマトンボ Epophthalmia elegans elegans など,大型で幼虫成長に時間がかかる種が含まれます.また一年目の冬を卵で越冬するルリボシヤンマ属のオオルリボシヤンマ Aeshna crenata やルリボシヤンマ Aeshna juncea juncea も多年一化の夏季種です.
Norling & Sahlén (1997)は,summer species という語を使わず,late breeding species (遅い時期に繁殖活動をする種)という語をあてています.そしてこれらの種について次のように述べています.「このグループの種は(生活史を)はっきりと特徴づけることができず,またその生物季節は緯度とともに変化する.温暖な季節が短くなるもっとも北の分布地域では,これらはすべて early breeders のようになる」.
これに関する例として Norling (1984) は,スェーデンの異なる緯度の生息地で,エゾイトトンボ属の Coenagrion hastulatum を使って,生活史のコントロールについて調べています.その中で,スェーデンの南北緯58度42分の生息地では一年一化のものと二年一化のものが混在し,羽化前の冬をF-0,F-1,F-2*3の幼虫3齢期で過ごしているのに対し,同じスェーデンの北緯67度50分の生息地では三年または四年一化の生活史となり,羽化前の冬をほとんど終齢幼虫で過ごしていた,と述べています.
つまり,南方では夏季種の特徴を有していたものが北方では春季種の特徴を有するようになっているということです.これは,夏季種の生活史の可塑性(化性を変更できること)を物語ると同時に,特に北方への分布拡大によって,夏季種から春季種へと生活史が進化していくことを暗示しているものとして,とても興味深い事実です.別の見方をすれば,春季種と夏季種の間には連続的な生活史の変異があって,明確に区別できない場合があるともいえるでしょう.
Ⅴ.秋季種の生活史
秋季種は原則として卵で越冬し,かつ一年一化であるという生活史を持つものとして定義されます(上田,1990).兵庫県に生息するトンボでは,ネキトンボ Sympetrum speciosum speciosum を除くアカネ属 Genus Sympetrum のトンボ14種,アオイトトンボ属 Genus Lestes の3種,ヤンマ科のカトリヤンマ Gynacantha japonica がこのカテゴリーに入ると考えられます*4.もう少し範囲を広げると,信州以北に分布するマダラヤンマ Aeshna mixta soneharai も卵で越冬し一年一化です.
秋季種は卵で越冬するので,必然的に卵で過ごす期間が長くなります.常温での飼育卵期間が80日以上あるものが多く,これは Ando (1962) のいう diapause species,Corbet (1962) のいう delayed development eggs に該当します.つまり,卵発生が途中で停止するということです.また経験的にも記録においても,秋季種の幼虫がほとんど冬季に採集できないということから,実態として卵で越冬していることはほぼ間違いありません.
さらにまた,秋季種の幼虫が春から夏にかけてしか採集できないという経験的事実から,卵越冬を実現した秋季種は,春に孵化し,その後幼虫が急速に成長し,初夏に羽化するという事実もほぼ間違いのないことといえるでしょう.
しかし秋季種の卵や幼虫を飼育してみると,その結果にはかなりばらつきが見られます(例えば;吉田,2014).卵期間に,越冬期間の長さから見るとはるかに短い40-60日程度の結果が出たり,幼虫期間に,半年以上という,とても春に孵化したものが初夏には羽化できないような長さの結果が出たりしています.これらを考察する上では,飼育環境と実際の卵や幼虫の置かれている環境の違いを,十分に見ていかねばならないでしょう.
例えば,打空産卵をするアカネ属の飼育卵期間が短いという傾向があります.上田(1996)は,「水に浸かった状態では卵の休眠が生じないか,あるいは短くなってしまう種は,卵越冬を維持するためには卵を水のないところに置く必要があり,水のない場所に卵を産むことを可能にする打空産卵は,そのような性質の卵を持った種において有利な産卵方式であるに違いない」との考え方を述べています.
多年一化の種としては,オニヤンマ科のオニヤンマ Anotogaster sieboldii,サナエトンボ科のウチワヤンマ Sinictinogomphus clavatus clavatus,ヤンマ科のコシボソヤンマ Boyeria maclachlani やミルンヤンマ Planaeschna milnei milnei,エゾトンボ科のエゾトンボ Somatochlora viridiaenea やハネビロエゾトンボ Somatochlora clavata,ヤマトンボ科のオオヤマトンボ Epophthalmia elegans elegans など,大型で幼虫成長に時間がかかる種が含まれます.また一年目の冬を卵で越冬するルリボシヤンマ属のオオルリボシヤンマ Aeshna crenata やルリボシヤンマ Aeshna juncea juncea も多年一化の夏季種です.
Norling & Sahlén (1997)は,summer species という語を使わず,late breeding species (遅い時期に繁殖活動をする種)という語をあてています.そしてこれらの種について次のように述べています.「このグループの種は(生活史を)はっきりと特徴づけることができず,またその生物季節は緯度とともに変化する.温暖な季節が短くなるもっとも北の分布地域では,これらはすべて early breeders のようになる」.
これに関する例として Norling (1984) は,スェーデンの異なる緯度の生息地で,エゾイトトンボ属の Coenagrion hastulatum を使って,生活史のコントロールについて調べています.その中で,スェーデンの南北緯58度42分の生息地では一年一化のものと二年一化のものが混在し,羽化前の冬をF-0,F-1,F-2*3の幼虫3齢期で過ごしているのに対し,同じスェーデンの北緯67度50分の生息地では三年または四年一化の生活史となり,羽化前の冬をほとんど終齢幼虫で過ごしていた,と述べています.
つまり,南方では夏季種の特徴を有していたものが北方では春季種の特徴を有するようになっているということです.これは,夏季種の生活史の可塑性(化性を変更できること)を物語ると同時に,特に北方への分布拡大によって,夏季種から春季種へと生活史が進化していくことを暗示しているものとして,とても興味深い事実です.別の見方をすれば,春季種と夏季種の間には連続的な生活史の変異があって,明確に区別できない場合があるともいえるでしょう.
Ⅴ.秋季種の生活史
秋季種は原則として卵で越冬し,かつ一年一化であるという生活史を持つものとして定義されます(上田,1990).兵庫県に生息するトンボでは,ネキトンボ Sympetrum speciosum speciosum を除くアカネ属 Genus Sympetrum のトンボ14種,アオイトトンボ属 Genus Lestes の3種,ヤンマ科のカトリヤンマ Gynacantha japonica がこのカテゴリーに入ると考えられます*4.もう少し範囲を広げると,信州以北に分布するマダラヤンマ Aeshna mixta soneharai も卵で越冬し一年一化です.
秋季種は卵で越冬するので,必然的に卵で過ごす期間が長くなります.常温での飼育卵期間が80日以上あるものが多く,これは Ando (1962) のいう diapause species,Corbet (1962) のいう delayed development eggs に該当します.つまり,卵発生が途中で停止するということです.また経験的にも記録においても,秋季種の幼虫がほとんど冬季に採集できないということから,実態として卵で越冬していることはほぼ間違いありません.
さらにまた,秋季種の幼虫が春から夏にかけてしか採集できないという経験的事実から,卵越冬を実現した秋季種は,春に孵化し,その後幼虫が急速に成長し,初夏に羽化するという事実もほぼ間違いのないことといえるでしょう.
しかし秋季種の卵や幼虫を飼育してみると,その結果にはかなりばらつきが見られます(例えば;吉田,2014).卵期間に,越冬期間の長さから見るとはるかに短い40-60日程度の結果が出たり,幼虫期間に,半年以上という,とても春に孵化したものが初夏には羽化できないような長さの結果が出たりしています.これらを考察する上では,飼育環境と実際の卵や幼虫の置かれている環境の違いを,十分に見ていかねばならないでしょう.
例えば,打空産卵をするアカネ属の飼育卵期間が短いという傾向があります.上田(1996)は,「水に浸かった状態では卵の休眠が生じないか,あるいは短くなってしまう種は,卵越冬を維持するためには卵を水のないところに置く必要があり,水のない場所に卵を産むことを可能にする打空産卵は,そのような性質の卵を持った種において有利な産卵方式であるに違いない」との考え方を述べています.
*3. FはFinalの略号で終齢を表します.「-数字」は終齢から逆算しての齢数を表します.つまり,F-0は終齢,F-1は亜終齢,F-2は終齢二つ手前ということです.
*4. 尾園ら(2012)には,ミヤマアカネについて,「一部は一年二化している可能性がある」との記述があります.これは夏早くや秋遅くの未熟成虫の出現や,卵期間・幼虫期間の最短記録から推定される可能性で,幼虫の成育を野外で追跡したものではないようです.しかし,今後検討すべき面白い状況ではあります.

写真7.秋季種と考えられるミヤマアカネ(左:2009.9.13.)とマユタテアカネ(右:2009.8.30.)の遅い羽化.右下は同日の産卵.
秋季種は,特に温暖な地域において,成虫が生殖休眠を行います(例えば;上田,1990).つまり羽化した後,だいたい1~2週間程度とされる通常の前生殖期をはるかに超える期間,生殖活動が始まらないということです.兵庫県では,秋季種の多くは6-7月頃に羽化していますが,この生殖休眠によって繁殖活動が秋まで遅延されることになるわけです.したがって,成虫の出現期間は数ヶ月と,一般に長くなります.
このうち前生殖期の長さについてはアオイトトンボ Lestes sponsa で詳しく研究がなされていて,これに地理的クライン*5が存在することが知られています.それによると,緯度が北になるほど,前生殖期*6の長さが短くなり,北緯40度を超えたあたりからはおよそ20日間で一定値になります(Uéda,1978).この結果は,秋季種でも,夏が短く,前生殖期を延長しなくても産卵時期の秋がやって来るような北の地域では,生殖休眠がなくなる場合があるということです.つまり南の地域での生殖休眠は卵越冬を実現するために秋まで繁殖期を遅らせる生活史戦略と考えることができます(上田,1990).
以上のように,秋季種は,卵越冬し,春に孵化し,幼虫の発育速度が速く,初夏に羽化し,温暖な地域では長い前生殖期間を過ごし,秋に繁殖活動を行う種と一般化することができます.
このうち前生殖期の長さについてはアオイトトンボ Lestes sponsa で詳しく研究がなされていて,これに地理的クライン*5が存在することが知られています.それによると,緯度が北になるほど,前生殖期*6の長さが短くなり,北緯40度を超えたあたりからはおよそ20日間で一定値になります(Uéda,1978).この結果は,秋季種でも,夏が短く,前生殖期を延長しなくても産卵時期の秋がやって来るような北の地域では,生殖休眠がなくなる場合があるということです.つまり南の地域での生殖休眠は卵越冬を実現するために秋まで繁殖期を遅らせる生活史戦略と考えることができます(上田,1990).
以上のように,秋季種は,卵越冬し,春に孵化し,幼虫の発育速度が速く,初夏に羽化し,温暖な地域では長い前生殖期間を過ごし,秋に繁殖活動を行う種と一般化することができます.
*5.クラインとはある方向に沿って連続的に変化する形質をさす術語です.
*6. この場合は(個体群の)最初の羽化から最初の産卵までの期間をさしています.
トンボの生態学 7.生活史の季節的制御
トンボの休眠
生活史の季節的制御において,その中心的枠割りを果たすものとして「休眠」という現象があります.このことは正木進三博士の著作「昆虫の生活史の進化」を読めば一目瞭然でしょう.そこでここでは,トンボにおける休眠現象について解説を試みたいと思っています.「試みる」というやや消極的な表現になっているのは,実際の休眠現象はその解釈に非常に難しい側面があるからです.前節で生活史の説明をするときにも,私は「休眠」という語をできるだけ使わないようにしました.しかし,生活史の季節的制御について論じるときに「休眠」は避けることのできない現象ですから,自分なりの解釈を交えて,ここにまとめておきたいと思います.
Ⅰ.広義の休眠,休眠,休止,そして休眠発育
休眠という語を岩波の生物学辞典(第四版)で引いてみますと,大きく二つに分けて解説がなされています.その一つには dormancy という語,もう一つには diapause という語があてられています.つまり異なる意味のものが,英語ではきちんと二つの語で使い分けられているのに対し,日本語では一つの「休眠」という語で表現されていることになります.そこで,本ページでは,以後これを区別するために,dormancy には広義の休眠,diapause に休眠という語をあてることにしました.
生物学辞典によりますと,dormancy は「動物の発生過程に起きる成長や活動の一時的停止の全現象を含む」とあり,diapause は「低温など特殊な環境条件下に置かれたり一定期間経過しないと,活動が再開されないような休眠」とあります.つまり,diapause は dormancy の中のある特殊な状態のものをさしているということです.
さらに dormancy の解説中に,休止 quiescence という語が出てきます.これは,大雑把に言えば,dormancy のうち diapause を除いた部分を示す語で,発生や活動の一時的停止を引き起こしている悪条件が取り除かれれば,ただちに発生や活動が再開されるような状態のことを意味します.対して,diapause では,こういった悪条件が取り除かれても,ある環境の合図を受けるまで,または一定期間経過しないと,発生や活動が再開されない,あるいは,発育に悪条件でないと思われる環境条件下でも,何らかの環境の合図で発育が停止したり再開されたりする,ということになります.そして発生や活動が再開したとき,diapause は消去されたといいます.
さらに休眠 diapause は,その誘導のされ方の違いによって,内因性休眠(または絶対的休眠) obligate diapause と外因性休眠(または条件的休眠) facultative diapause に分けられます(英術語はCorbet, 1999に従った).前者は,発生の過程で遺伝的に決められたステージで,環境条件のいかんに関わらず誘導される休眠で,後者は,ある環境条件になったとき,その合図 (cue) を受け取って休眠が誘導される場合をいいます.繰り返しになりますが,いずれの場合も,休眠の消去には,ある環境条件の実現,または一定期間の経過が不可欠です.
野外で見られる昆虫などの広義の休眠現象は,この休眠と休止が入り交じって現れてきているのが一般的で,概念的には休眠と休止が分離できたとしても,現実の状態にそれを区別して当てはめるのは困難な場合が多いことは知っておく必要があります.
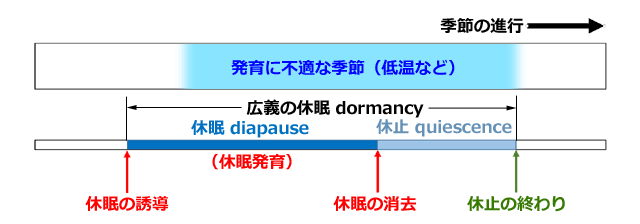
さて,もう一つ,休眠について知っておくべき術語があります.それは休眠発育 diapause development です.休眠 diapause 中は,形態形成や活動が停止しているように見えますが,実は次の段階に進むための生理的な変化が体内で進行しているのです.この時期の生理的変化を指す語が「休眠発育」です.休眠発育が完了すると次のステップに進むことができるため,休眠発育の生理は休眠の覚醒機構と深く結びついていると考えられています(例えば,園部,1990).したがって,休眠発育を行っている期間が休眠の期間であるともいえます (Corbet, 1999).
Ⅱ.卵の休眠
卵に発育遅延が生じた証拠を,通常の飼育卵期間が80日以上 (delayed development eggs) になる場合があるものとすると,吉田(2014)が総括した卵期間の一覧表からみて,日本では,卵越冬する秋季種のトンボと,産卵に続く冬を卵で越す多年一化の夏季種にだけ発育遅延が見られるといってよいでしょう.
卵のステージに休眠が組み込まれているかどうかは,卵期間だけからははっきりとさせることができません.卵休眠とは卵内で発生している胚の発育停止のことですから,胚発生を継続的に観察しなければ,発育停止の有無を捕らえることができません.胚の形態形成にまで注目してトンボの卵の休眠現象を捉えた研究はそれほど多くありませんが,その中でよく引用されているものに,安藤裕博士の有名な研究があります (Ando,1962).
これによると,トンボの胚発生が停止するステージには大きく二つあって,一つは胚反転 katatrepsis*7 の起きる前のステージ,もう一つは胚反転を終え胚発生がほぼ完了したステージです.前者では,ルリボシヤンマとオオルリボシヤンマ (Aeshna-type),後者では,アオイトトンボとアキアカネ (それぞれ Lestes-,Sympetrum- types) が該当します.研究に用いたこれら卵越冬性のトンボについては,観察温度条件が記載されていないため,この結果だけからは,これらの発育停止が休眠か休止かは判断しにくい部分があります.
Ⅰ.広義の休眠,休眠,休止,そして休眠発育
休眠という語を岩波の生物学辞典(第四版)で引いてみますと,大きく二つに分けて解説がなされています.その一つには dormancy という語,もう一つには diapause という語があてられています.つまり異なる意味のものが,英語ではきちんと二つの語で使い分けられているのに対し,日本語では一つの「休眠」という語で表現されていることになります.そこで,本ページでは,以後これを区別するために,dormancy には広義の休眠,diapause に休眠という語をあてることにしました.
生物学辞典によりますと,dormancy は「動物の発生過程に起きる成長や活動の一時的停止の全現象を含む」とあり,diapause は「低温など特殊な環境条件下に置かれたり一定期間経過しないと,活動が再開されないような休眠」とあります.つまり,diapause は dormancy の中のある特殊な状態のものをさしているということです.
さらに dormancy の解説中に,休止 quiescence という語が出てきます.これは,大雑把に言えば,dormancy のうち diapause を除いた部分を示す語で,発生や活動の一時的停止を引き起こしている悪条件が取り除かれれば,ただちに発生や活動が再開されるような状態のことを意味します.対して,diapause では,こういった悪条件が取り除かれても,ある環境の合図を受けるまで,または一定期間経過しないと,発生や活動が再開されない,あるいは,発育に悪条件でないと思われる環境条件下でも,何らかの環境の合図で発育が停止したり再開されたりする,ということになります.そして発生や活動が再開したとき,diapause は消去されたといいます.
さらに休眠 diapause は,その誘導のされ方の違いによって,内因性休眠(または絶対的休眠) obligate diapause と外因性休眠(または条件的休眠) facultative diapause に分けられます(英術語はCorbet, 1999に従った).前者は,発生の過程で遺伝的に決められたステージで,環境条件のいかんに関わらず誘導される休眠で,後者は,ある環境条件になったとき,その合図 (cue) を受け取って休眠が誘導される場合をいいます.繰り返しになりますが,いずれの場合も,休眠の消去には,ある環境条件の実現,または一定期間の経過が不可欠です.
野外で見られる昆虫などの広義の休眠現象は,この休眠と休止が入り交じって現れてきているのが一般的で,概念的には休眠と休止が分離できたとしても,現実の状態にそれを区別して当てはめるのは困難な場合が多いことは知っておく必要があります.
図2.野外で休眠から休止へ移行する例の概念図.冬に休眠する例で示した.
まだ発育に不適でない時期に遺伝的プログラムまたは環境の合図で休眠が誘導される.
休眠発育を終え,低温など環境の合図で休眠が消去される.
しかしそのときは発育に不適切な温度のため発育が停止状態(=休止)となる.
そして発育に適切な環境になったときに休止が終了し,発育が再開される.
しかし休眠から休止への移行時期についてははっきりさせることが困難である.
しかしながら,休眠と休止を区別できる場合もあります.通常昆虫の発育速度は,生育に適した温度範囲内では,温度と正の相関関係があります.つまり温度が高くなると速く発育するということです.これはよく知られている有効積算温度の法則として一般化されています.しかし休眠が生じている場合には,この相関関係が成り立たなくなったり,逆に負の相関関係 (*Boehms,1971) になったりします.そしてこのことを拡張して考えると,発育に適した環境条件でありながら発育に停止や遅延が生じている場合,休眠 diapause が起きていると判断できます.同様のことは生物学辞典にも記述があります.まだ発育に不適でない時期に遺伝的プログラムまたは環境の合図で休眠が誘導される.
休眠発育を終え,低温など環境の合図で休眠が消去される.
しかしそのときは発育に不適切な温度のため発育が停止状態(=休止)となる.
そして発育に適切な環境になったときに休止が終了し,発育が再開される.
しかし休眠から休止への移行時期についてははっきりさせることが困難である.
さて,もう一つ,休眠について知っておくべき術語があります.それは休眠発育 diapause development です.休眠 diapause 中は,形態形成や活動が停止しているように見えますが,実は次の段階に進むための生理的な変化が体内で進行しているのです.この時期の生理的変化を指す語が「休眠発育」です.休眠発育が完了すると次のステップに進むことができるため,休眠発育の生理は休眠の覚醒機構と深く結びついていると考えられています(例えば,園部,1990).したがって,休眠発育を行っている期間が休眠の期間であるともいえます (Corbet, 1999).
Ⅱ.卵の休眠
卵に発育遅延が生じた証拠を,通常の飼育卵期間が80日以上 (delayed development eggs) になる場合があるものとすると,吉田(2014)が総括した卵期間の一覧表からみて,日本では,卵越冬する秋季種のトンボと,産卵に続く冬を卵で越す多年一化の夏季種にだけ発育遅延が見られるといってよいでしょう.
卵のステージに休眠が組み込まれているかどうかは,卵期間だけからははっきりとさせることができません.卵休眠とは卵内で発生している胚の発育停止のことですから,胚発生を継続的に観察しなければ,発育停止の有無を捕らえることができません.胚の形態形成にまで注目してトンボの卵の休眠現象を捉えた研究はそれほど多くありませんが,その中でよく引用されているものに,安藤裕博士の有名な研究があります (Ando,1962).
これによると,トンボの胚発生が停止するステージには大きく二つあって,一つは胚反転 katatrepsis*7 の起きる前のステージ,もう一つは胚反転を終え胚発生がほぼ完了したステージです.前者では,ルリボシヤンマとオオルリボシヤンマ (Aeshna-type),後者では,アオイトトンボとアキアカネ (それぞれ Lestes-,Sympetrum- types) が該当します.研究に用いたこれら卵越冬性のトンボについては,観察温度条件が記載されていないため,この結果だけからは,これらの発育停止が休眠か休止かは判断しにくい部分があります.

写真8.アオイトトンボ.左は羽化(6月21日),右はハリイの一種に産卵(9月14日)
このうちアオイトトンボに関しては,イギリスで行われた Corbet (1956) の古典的研究がありますのでこれを紹介しましょう.7月1日にハリイ属の一種にアオイトトンボが産卵していました.その茎に印をつけそのまま野外に置き7月17日に採取,8月13日まで27日間20℃で保存しました.そしてこのうち20卵を取り出して観察したところ,胚運動 blastokinesis はすべて完了していました.この場合の胚運動は胚反転のことだと思われます.またこの胚運動の完了までの2週間,遅延なく発生が進むことが過去の研究から知られていますので,7月1日から17日の間に新たに産卵された卵があったとしても,この27日間の間にすべて胚運動を完了している計算になります(右図).
この胚運動が完了した卵を茎から取り出して水に浸し,いくつかの異なる処理を行いました.一つは20℃にずっと置き,あとは,3,6,9,12,15週間それぞれ10℃及び5℃に置いた後20℃に戻し,孵化までの日数を調べました.なお,10℃,5℃というのは孵化が起きる閾値より低い温度で,20℃はそれを超えています.結果は,20℃に戻してから孵化するまでの日数で,大きく二つのグループに分けられました.一つは孵化に90-217日かかったグループA(図4黒色),もう一つは5-28日のうちに孵化が起きたグループB(図4赤色)です.
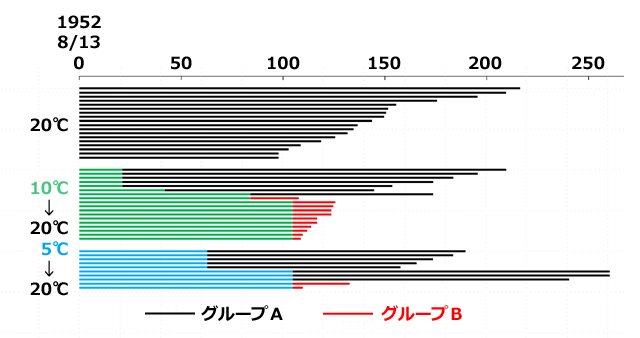
一方10℃に15週間置いた実験群では,20℃に戻したときにいずれも短期間で孵化が生じています.これは,休眠発育が10℃で15週間以内に完了し,20℃に戻したときに孵化に向かって発生が急速に進行したためと解釈できます.対して,10℃12週間や5℃15週間の実験群では,休眠発育が完了していたりしなかったりするという結果になっています.つまり休眠発育は,10℃12週間では期間がぎりぎり短く,5℃ではその速度が遅くなるということでしょう.
ところが,図4から次のような疑問が浮かびます.休眠発育の完了には適度な低温に適度な期間さらされるという条件が重要なようだが,どの実験群もトータルで90-250日以上かかって孵化しているわけなので,卵越冬するという点では,この条件にあまり重要な意味があるとは考えられないということです.秋に発育が遅延さえすれば,やがて温度は孵化の閾値以下となり,結局胚に休止が生じて,冬を越せることになるのではないか,という考えが成り立つということです.
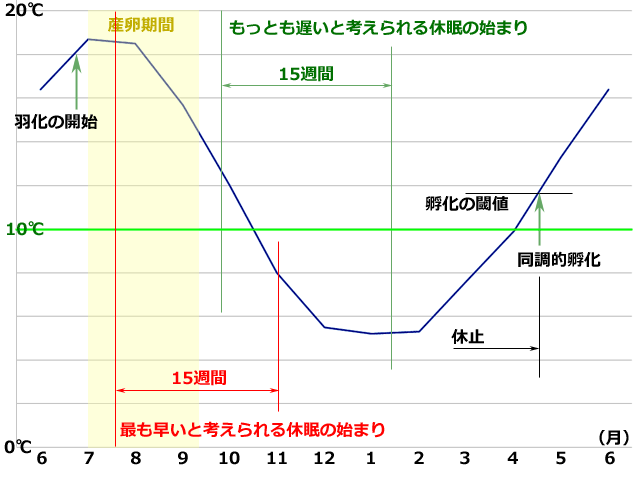
これに関して Corbet (1956) は,グループBの短期間に起きる孵化の同調性に着目して,低温にさらされる意味は,この同調性を高めることにあるとしています.夏の終わりに産下された卵は,孵化の閾値より高い夏の終わりから,秋・冬へと次第に気温が下がって孵化の閾値を下回るまで,休眠発育が継続します.そして5℃以下になる厳冬期までには休眠発育を終え発育がそろいます.そして早く休眠発育を終えたものから順次休止状態になっていき,春を待つことになります.やがて春が来て気温が孵化の閾値以上になったときに,発育のそろった卵が同調的に孵化する,という意味のことを述べています(図5参照).
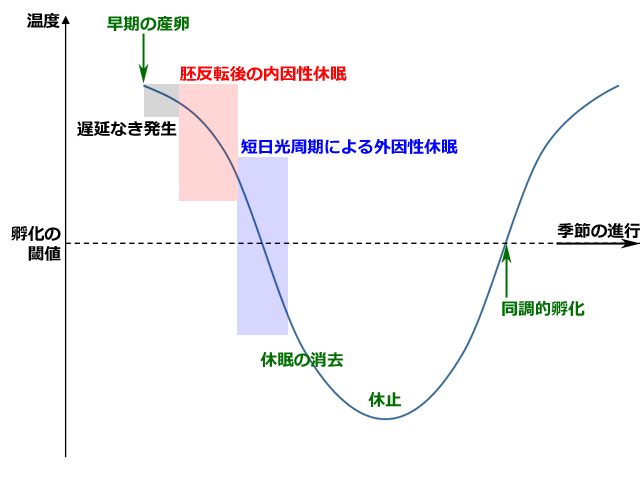
さて,早い時期に産下された卵では,胚反転が完了するまで遅延なく発生が進んでから,休眠に入ります.一方遅い時期に産下された卵では,胚反転の前までは遅延なく発生が進みますが,その時点で休眠に入ります.この胚の休眠発育には最適温度(10-14℃)があって,それより高い温度でも低い温度でも,休眠発育が長期化してしまいます.これによって休眠発育が孵化閾値より低い温度で完了します.
以上いくつかの研究を紹介しましたが,トンボの卵による休眠の意義は,以下のようにまとめられると思います.
二つ目は,幼虫の速い成長速度と相まって,ほぼ同じ時期に成虫を出現させることができる点で重要な意味を持ちます.特に北方の夏が短い地域では,早い時期の同調的孵化は,繁殖活動の期間を少しでも長く確保できるということにつながります.また,同調的に孵化することで生息地個体群全体の幼虫期間のばらつきが少なくなり,全体として自由水を必要とする時間が短くなることで,干上がりに対する適応が高くなるともいえるでしょう.
トンボの越冬卵は,低温処理すると孵化の同調性が高まるとよく言われてきましたが,これは低温処理によって休眠発育を終えさせ,さらに休止状態に置くことで,温度を上げたときに同調的に孵化できることによると考えられます.したがって,低温処理にはそれなりの長さが必要ということになります.
Ⅲ.幼虫の休眠
Corbet (1999) は,幼虫の休眠について,「... diapause can be averted by appropriate treatment; so, ..., it must be regarded as facultative rather than obligate. ([これまで調べられてきた幼虫の]休眠は適切な処理によって避けることができるので,内因性(絶対的)というよりは外因性(条件的)と見なすべきである)」と述べています.また,卵の休眠のところでも述べたように,休眠と休止を見分けることも困難であると述べています.
幼虫に知られている休眠と考えられる現象は,大きく二つあります.一つは夏における発育遅延または停滞と,冬の発育停止です.成長可能な暖かい季節に幼虫の発育が停滞したり遅延したりする現象は休眠が生じている可能性が高いと考えられますが,冬の場合は,休眠しないものでも,低温によって休止しますから,休眠が生じていることを示すためには,綿密な実験を計画する必要があります.
この点に関して,例えば Norling (1976) は,南スェーデン(北緯58度43分)に生息するカオジロトンボ原名亜種 Leucorrhinia dubia dubia を使って,緻密な実験を行っています.それによると,20℃の恒温飼育条件下で季節に関わりなく,F-4齢からF-1齢の幼虫に対し,短日光周期(LD13:11;明期13時間:暗期11時間)が,それらの齢期間の長期化を引き起こしたことを示しています.したがって,この発育遅延は休眠によるものと考えて問題ないと思われます.また休眠が短日という環境の合図によって引き起こされているので,これは外因性休眠であるといえます.この結果を自然条件に当てはめれば,夏の間成長を続けていた幼虫のうち,夏の終わりにF-4齢からF-1齢に達したものは,秋分の日あたりの日長条件(薄明薄暮を含めるとLD13:11ぐらいになる)で発育速度がぐんと落ちるということになります.
同じ論文で,彼は終齢幼虫について,特定の発育段階にある個体(複眼の発達で区別されたフェーズ2や3)が,長日(LD19:3他)によってそのステージが長期化すること,つまり長日休眠することも示しています.この研究では,この個体群の長日休眠の臨界日長がLD17.7:6.3付近にあることも示されていて,こういった光周性に関する緻密な研究は,トンボではあまり見かけることがありません.
この胚運動が完了した卵を茎から取り出して水に浸し,いくつかの異なる処理を行いました.一つは20℃にずっと置き,あとは,3,6,9,12,15週間それぞれ10℃及び5℃に置いた後20℃に戻し,孵化までの日数を調べました.なお,10℃,5℃というのは孵化が起きる閾値より低い温度で,20℃はそれを超えています.結果は,20℃に戻してから孵化するまでの日数で,大きく二つのグループに分けられました.一つは孵化に90-217日かかったグループA(図4黒色),もう一つは5-28日のうちに孵化が起きたグループB(図4赤色)です.
図4.イギリスのアオイトトンボの孵化までの期間 (Corbet, 1956をもとに描画).
緑色・青色はそれぞれ10℃,5℃に置いた期間を表す.
黒色・赤色はそれぞれ20℃に戻して孵化するまでの期間を表す.
実験開始時期には,形態的な胚発生自体はほぼ完了していることに注意すること.
したがってこれは形態的な胚発生完了後孵化までにどれくらいの時間がかかるかを示している.
処理方法によって,20℃に戻してから孵化までが長いグループAと短いグループBに分かれる.
この結果からいくつかのことが結論づけられます.まず孵化の温度閾値を越えている20℃の実験群で,形態形成が完了した胚の孵化が90日以上遅延されることから,アオイトトンボの卵に休眠 (内因性休眠;Corbet, 1956) が生じていることはまず間違いないということです.そしてこの休眠中に生じている休眠発育が,20℃のままでは非常に時間がかかる結果,孵化の遅延が生じていると解釈できることです.緑色・青色はそれぞれ10℃,5℃に置いた期間を表す.
黒色・赤色はそれぞれ20℃に戻して孵化するまでの期間を表す.
実験開始時期には,形態的な胚発生自体はほぼ完了していることに注意すること.
したがってこれは形態的な胚発生完了後孵化までにどれくらいの時間がかかるかを示している.
処理方法によって,20℃に戻してから孵化までが長いグループAと短いグループBに分かれる.
一方10℃に15週間置いた実験群では,20℃に戻したときにいずれも短期間で孵化が生じています.これは,休眠発育が10℃で15週間以内に完了し,20℃に戻したときに孵化に向かって発生が急速に進行したためと解釈できます.対して,10℃12週間や5℃15週間の実験群では,休眠発育が完了していたりしなかったりするという結果になっています.つまり休眠発育は,10℃12週間では期間がぎりぎり短く,5℃ではその速度が遅くなるということでしょう.
ところが,図4から次のような疑問が浮かびます.休眠発育の完了には適度な低温に適度な期間さらされるという条件が重要なようだが,どの実験群もトータルで90-250日以上かかって孵化しているわけなので,卵越冬するという点では,この条件にあまり重要な意味があるとは考えられないということです.秋に発育が遅延さえすれば,やがて温度は孵化の閾値以下となり,結局胚に休止が生じて,冬を越せることになるのではないか,という考えが成り立つということです.
これに関して Corbet (1956) は,グループBの短期間に起きる孵化の同調性に着目して,低温にさらされる意味は,この同調性を高めることにあるとしています.夏の終わりに産下された卵は,孵化の閾値より高い夏の終わりから,秋・冬へと次第に気温が下がって孵化の閾値を下回るまで,休眠発育が継続します.そして5℃以下になる厳冬期までには休眠発育を終え発育がそろいます.そして早く休眠発育を終えたものから順次休止状態になっていき,春を待つことになります.やがて春が来て気温が孵化の閾値以上になったときに,発育のそろった卵が同調的に孵化する,という意味のことを述べています(図5参照).
図5.この実験結果を自然状態に当てはめてみたもの (Corbet, 1956をもとに描画).
折れ線グラフは,観察地(ウォークフィールド・コモン)とほぼ同緯度のロンドンの平均気温
(1981-2010;https://www.travel-zentech.jp/world/infomation/kion/uk.htm より)
羽化,産卵期間,孵化の各時期は Corbet (1956) の記述による.
卵の産下から2週間で胚反転を完了し休眠に入ると仮定して休眠の始まりを決めた.
15週間は一つの目安として休眠発育の最短期間(10℃15週間)を示している.
実際は10℃より温度が高かったり低かったりするので,より長い間かかっているはずだ.
しかしいずれの場合も厳冬期が終わるまでには,休眠発育を終えることができそうである.
カナダのサスカチワン州(北緯52度)で調べられた,同じアオイトトンボ属の Lestes disjuncus も似たような休眠を行って,卵越冬に入ります.この休眠(内因性休眠)は胚反転を終えてから誘起されますが,早い時期に産下された卵は,まだ卵の孵化閾値を上回っている間に休眠発育が完了してしまいます.ところがこの卵には,秋の短日光周期(12-14明期)に誘導される外因性休眠が,引き続き生じるのです.これによって,内因性休眠の終了が孵化閾値以上の温度であっても,秋の孵化が抑制されます.また遅く産下された卵は,温度が孵化閾値以下になるまで,内因性休眠が続くことになります.いずれにしても,これらの休眠は冬の間に消去され,低温による休止を経て,春に同調的な孵化が生じます(以上図6,*Sawchyn & Gillott, 1974;*Sawchyn & Church, 1973).折れ線グラフは,観察地(ウォークフィールド・コモン)とほぼ同緯度のロンドンの平均気温
(1981-2010;https://www.travel-zentech.jp/world/infomation/kion/uk.htm より)
羽化,産卵期間,孵化の各時期は Corbet (1956) の記述による.
卵の産下から2週間で胚反転を完了し休眠に入ると仮定して休眠の始まりを決めた.
15週間は一つの目安として休眠発育の最短期間(10℃15週間)を示している.
実際は10℃より温度が高かったり低かったりするので,より長い間かかっているはずだ.
しかしいずれの場合も厳冬期が終わるまでには,休眠発育を終えることができそうである.
図6.早く産下された Lestes disjuncus の卵における休眠と卵越冬の概念図.
曲線は温度変化を模式的に表している.
外因性休眠の消去時期についてははっきりしていない.
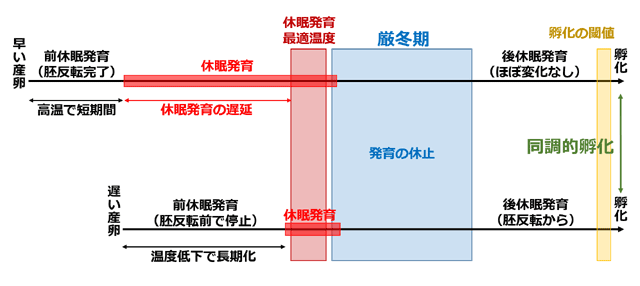
またアカネ属の Sympetrum vicinum でも,アオイトトンボと似たような休眠が知られています(*Boehum, 1971;北緯37度,ノースカロライナ州).違うところは,休眠に入る胚発生のステージが,早く産下された卵と遅く産下された卵で異なっているという点です.これに関して Corbet (1999) は「embryonic stage is not a reliable indicator of physiological stage (胚発生の(形態的)ステージは,生理的ステージの信頼性のある指標ではない)」と述べています.曲線は温度変化を模式的に表している.
外因性休眠の消去時期についてははっきりしていない.
さて,早い時期に産下された卵では,胚反転が完了するまで遅延なく発生が進んでから,休眠に入ります.一方遅い時期に産下された卵では,胚反転の前までは遅延なく発生が進みますが,その時点で休眠に入ります.この胚の休眠発育には最適温度(10-14℃)があって,それより高い温度でも低い温度でも,休眠発育が長期化してしまいます.これによって休眠発育が孵化閾値より低い温度で完了します.
図7.S. vicinum の卵における休眠発育と卵越冬の概念図 (*Boehum, 1971に基づく).
前休眠発育,休眠発育,後休眠発育は,「生理的」に分けられたステージで,
発生における形態的ステージと一致しないことに注意すること.
休眠発育期は,発生速度が温度と正の相関関係にならない時期として定義されている.
早い産下卵は休眠発育が大幅に遅延され,温度が孵化閾値以下になるまで休眠が消去されない.
原著論文が入手困難で,一部筆者の推定を含めている.
この通りになるには,後休眠発育の温度閾値が孵化の閾値より低いことが必要である.
上田(1993)は,アキアカネ Sympetrum frequens の卵休眠のステージが,定温の飼育温度によって変わってくるという実験結果を報告しています.アキアカネの卵を11℃や18℃で飼育すると,ステージ5(眼点や付属肢が形成されているのでおそらくほぼ胚発生を完了した時点)で発生が停止し,それぞれ80日またはさらに長くその状態が続きます.一方で25℃で飼育すると,ステージ2(顕微鏡下でようやく胚が確認できるので胚反転の前)で約40日間発生が停止します.この発育停止は休眠だと考えられますが,異なる形態的ステージで,生理的ステージである休眠が生じています.ただ温度が高い方が若い形態的ステージで休眠に入るという点で,Sympetrum vicinum とは逆になっています.前休眠発育,休眠発育,後休眠発育は,「生理的」に分けられたステージで,
発生における形態的ステージと一致しないことに注意すること.
休眠発育期は,発生速度が温度と正の相関関係にならない時期として定義されている.
早い産下卵は休眠発育が大幅に遅延され,温度が孵化閾値以下になるまで休眠が消去されない.
原著論文が入手困難で,一部筆者の推定を含めている.
この通りになるには,後休眠発育の温度閾値が孵化の閾値より低いことが必要である.
以上いくつかの研究を紹介しましたが,トンボの卵による休眠の意義は,以下のようにまとめられると思います.
- 暖かい秋に孵化してしまうことを避け,確実に卵越冬を実現させる.
- 春に同調的な孵化を実現する.
二つ目は,幼虫の速い成長速度と相まって,ほぼ同じ時期に成虫を出現させることができる点で重要な意味を持ちます.特に北方の夏が短い地域では,早い時期の同調的孵化は,繁殖活動の期間を少しでも長く確保できるということにつながります.また,同調的に孵化することで生息地個体群全体の幼虫期間のばらつきが少なくなり,全体として自由水を必要とする時間が短くなることで,干上がりに対する適応が高くなるともいえるでしょう.
トンボの越冬卵は,低温処理すると孵化の同調性が高まるとよく言われてきましたが,これは低温処理によって休眠発育を終えさせ,さらに休止状態に置くことで,温度を上げたときに同調的に孵化できることによると考えられます.したがって,低温処理にはそれなりの長さが必要ということになります.
Ⅲ.幼虫の休眠
Corbet (1999) は,幼虫の休眠について,「... diapause can be averted by appropriate treatment; so, ..., it must be regarded as facultative rather than obligate. ([これまで調べられてきた幼虫の]休眠は適切な処理によって避けることができるので,内因性(絶対的)というよりは外因性(条件的)と見なすべきである)」と述べています.また,卵の休眠のところでも述べたように,休眠と休止を見分けることも困難であると述べています.
幼虫に知られている休眠と考えられる現象は,大きく二つあります.一つは夏における発育遅延または停滞と,冬の発育停止です.成長可能な暖かい季節に幼虫の発育が停滞したり遅延したりする現象は休眠が生じている可能性が高いと考えられますが,冬の場合は,休眠しないものでも,低温によって休止しますから,休眠が生じていることを示すためには,綿密な実験を計画する必要があります.
この点に関して,例えば Norling (1976) は,南スェーデン(北緯58度43分)に生息するカオジロトンボ原名亜種 Leucorrhinia dubia dubia を使って,緻密な実験を行っています.それによると,20℃の恒温飼育条件下で季節に関わりなく,F-4齢からF-1齢の幼虫に対し,短日光周期(LD13:11;明期13時間:暗期11時間)が,それらの齢期間の長期化を引き起こしたことを示しています.したがって,この発育遅延は休眠によるものと考えて問題ないと思われます.また休眠が短日という環境の合図によって引き起こされているので,これは外因性休眠であるといえます.この結果を自然条件に当てはめれば,夏の間成長を続けていた幼虫のうち,夏の終わりにF-4齢からF-1齢に達したものは,秋分の日あたりの日長条件(薄明薄暮を含めるとLD13:11ぐらいになる)で発育速度がぐんと落ちるということになります.
同じ論文で,彼は終齢幼虫について,特定の発育段階にある個体(複眼の発達で区別されたフェーズ2や3)が,長日(LD19:3他)によってそのステージが長期化すること,つまり長日休眠することも示しています.この研究では,この個体群の長日休眠の臨界日長がLD17.7:6.3付近にあることも示されていて,こういった光周性に関する緻密な研究は,トンボではあまり見かけることがありません.
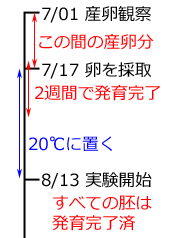

写真9.日本産カオジロトンボの成虫と幼虫.日本産のカオジロトンボは,亜種 Leucorrhinia dubia orientalis とされている.
これ以外にもいくつかの研究者による同様の研究報告があります.彼はそういった研究をレビューし,総説を書いています(Norling, 1984a).
休眠というと,生存に不適な季節を乗り越えるために行われるもの,と一般に思われているかも知れませんが,それだと,例えば暖かい季節に休眠することの意味が説明できません.実は,休眠は,成虫の繁殖期を特定の季節に合わせる機能も持っています.休眠によって,成虫の出現時期が調整されているとするならば,長期間幼虫で過ごし,かつ成虫出現期が限られるトンボほど,その調整のあり方が厳密になっていると考えられます.幼虫期間が長ければ温度条件や餌条件によって成長にばらつきが生じる可能性が高くなりますから,成虫の出現期間を限られたものにするには,その成長をどこかであるステージに収束させ,羽化を短期間で終わらせるしくみが必要になるはずです.このような例として,兵庫県南部(北緯35℃近く)のキイロサナエ Asiagomphus pryeri の幼虫休眠を紹介しましょう(以下 Aoki, 1999).
休眠というと,生存に不適な季節を乗り越えるために行われるもの,と一般に思われているかも知れませんが,それだと,例えば暖かい季節に休眠することの意味が説明できません.実は,休眠は,成虫の繁殖期を特定の季節に合わせる機能も持っています.休眠によって,成虫の出現時期が調整されているとするならば,長期間幼虫で過ごし,かつ成虫出現期が限られるトンボほど,その調整のあり方が厳密になっていると考えられます.幼虫期間が長ければ温度条件や餌条件によって成長にばらつきが生じる可能性が高くなりますから,成虫の出現期間を限られたものにするには,その成長をどこかであるステージに収束させ,羽化を短期間で終わらせるしくみが必要になるはずです.このような例として,兵庫県南部(北緯35℃近く)のキイロサナエ Asiagomphus pryeri の幼虫休眠を紹介しましょう(以下 Aoki, 1999).

写真10.キイロサナエの成虫と終齢幼虫.
キイロサナエは,多くが卵が産下されてから3回冬を越した後,3週間ほどで羽化します(図1).兵庫県では,羽化は5月中下旬から始まり,その後成虫は7月中頃まで,長くて約2ヶ月間見られます.繁殖活動は6月中にもっとも盛んになります.幼虫の成長を野外で追跡すると,羽化の前年から興味深い生育バターンが現れてきます.まず6月頃に羽化を終えた後,9月に入るまで終齢幼虫が出現しないのです.9月に入ると突然終齢幼虫が現れ,増えて,これらはそのまま越冬します.そして終齢で冬を越した個体だけが翌春温度依存的に遅延なく成長し羽化に至ります.つまり前年9月の同調的な終齢幼虫の出現が,翌年の同調的な羽化につながっているということです(図17参照).
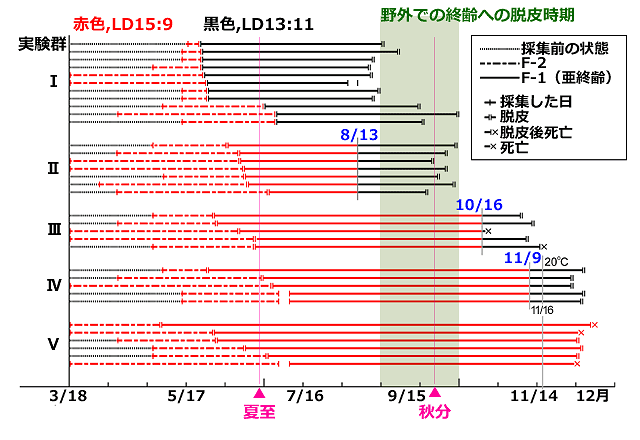
この実態は,夏の間は終齢幼虫への脱皮が抑制され9月にそれが解除される,というしくみを想定すれば説明ができます.上記のカオジロトンボ原名亜種の例から考えると,夏の長日休眠によって亜終齢(F-1)幼虫の脱皮が遅延され,秋の中間日長によってそれが消去されるとすれば説明できます.そこで,3-5月にかけてF-2齢幼虫を捕らえ,長日条件(夏至のころの日長 LD15:9)のもとで飼育し,亜終齢(F-1)に脱皮した後5つのグループに分け,様々な時期に中間日長(秋分のころの日長 LD13:11)に移し替えました.そして終齢への脱皮がいつ頃になるかを実験しました(図8).
これを自然状態に当てはめると,次のような発育の過程が導かれるでしょう.夏至前後にF-1齢に脱皮した幼虫は,そのころの長日よって休眠が誘導されて発育が遅延し,かつ発育の同調性が高められながら夏を過ごします.日長はだんだんと短くなっていき,やがて臨界日長(日長は不明だがおそらく秋分より前)を超えて,休眠が消去され,9月に同調的に終齢へと脱皮することとなります.そしてこの同調性が翌年の羽化が短期間で終了することにつながることになります.これを例え話でいうと,高速道路で同じインターを異なる時間にスタートした自動車が,渋滞によって次々追いつき,目的のインターにほぼ同時に着くことに似ています.
ところで,光周反応の実験は,上記のような固定日長で行われることが多いのですが,Corbet (1955) は,Anax imperator を用いた研究から,日長の絶対長ではなく変化する日長が,休眠の誘導にとって重要であると述べました.
この実態は,夏の間は終齢幼虫への脱皮が抑制され9月にそれが解除される,というしくみを想定すれば説明ができます.上記のカオジロトンボ原名亜種の例から考えると,夏の長日休眠によって亜終齢(F-1)幼虫の脱皮が遅延され,秋の中間日長によってそれが消去されるとすれば説明できます.そこで,3-5月にかけてF-2齢幼虫を捕らえ,長日条件(夏至のころの日長 LD15:9)のもとで飼育し,亜終齢(F-1)に脱皮した後5つのグループに分け,様々な時期に中間日長(秋分のころの日長 LD13:11)に移し替えました.そして終齢への脱皮がいつ頃になるかを実験しました(図8).
図8.キイロサナエ亜終齢(F-1)幼虫の長日休眠 (Aoki, 1999を改変).
F-2齢幼虫は3月18日から5月20日頃までに収集し,そのままLD15:9で飼育した.
実験群IはF-1齢に脱皮した日にLD13:11に移して飼育を継続した.
実験群II,III,IVは,それぞれ,8月13日,10月16日,11月9日に,LD13:11に移した.
実験群Vは,最後までLD15:9において飼育した.
温度は自然の推移のままで,11月16日以降は20℃を下回らないようにヒーターを投入した.
実験群Iの結果は,早くF-1に脱皮したものは早い時期に,遅くF-1に脱皮したものは遅く,それぞれ終齢への脱皮が起きています.対して,実験群Ⅱ,Ⅲ,Ⅳでは,いずれもLD13:11に移すまで終齢への脱皮が生じず,かつ,F-2からF-1への脱皮の時期にかかわらず,同調的に終齢に脱皮する傾向が強くなっていることが見て取れます.特に実験群Ⅲ,Ⅳでは,LD13:11への移行後,かなり短期間で終齢への脱皮が起きています.以上から,長日がF-1齢幼虫の休眠を誘導して発育を遅延させ,秋分付近の日長がそれを消去して終齢への脱皮を引き起こしているといえるでしょう.しかもこの長日休眠は終齢への脱皮の同調性を高める効果があることが分かります.F-2齢幼虫は3月18日から5月20日頃までに収集し,そのままLD15:9で飼育した.
実験群IはF-1齢に脱皮した日にLD13:11に移して飼育を継続した.
実験群II,III,IVは,それぞれ,8月13日,10月16日,11月9日に,LD13:11に移した.
実験群Vは,最後までLD15:9において飼育した.
温度は自然の推移のままで,11月16日以降は20℃を下回らないようにヒーターを投入した.
これを自然状態に当てはめると,次のような発育の過程が導かれるでしょう.夏至前後にF-1齢に脱皮した幼虫は,そのころの長日よって休眠が誘導されて発育が遅延し,かつ発育の同調性が高められながら夏を過ごします.日長はだんだんと短くなっていき,やがて臨界日長(日長は不明だがおそらく秋分より前)を超えて,休眠が消去され,9月に同調的に終齢へと脱皮することとなります.そしてこの同調性が翌年の羽化が短期間で終了することにつながることになります.これを例え話でいうと,高速道路で同じインターを異なる時間にスタートした自動車が,渋滞によって次々追いつき,目的のインターにほぼ同時に着くことに似ています.
ところで,光周反応の実験は,上記のような固定日長で行われることが多いのですが,Corbet (1955) は,Anax imperator を用いた研究から,日長の絶対長ではなく変化する日長が,休眠の誘導にとって重要であると述べました.
図1(再掲).キイロサナエの羽化曲線(Aoki, 1999)

写真11.Anax imperator (皇帝ギンヤンマ)のオス(左)とメス(右).
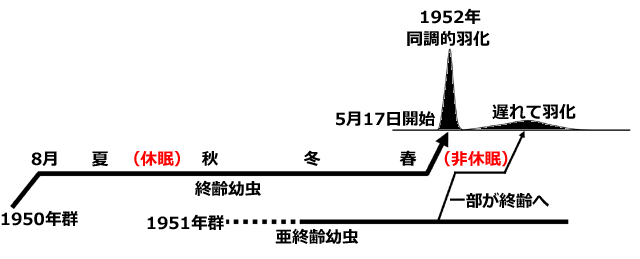
Corbet (1955) によると,Anax imperator の幼虫の羽化に至る付近の成長は次のようになっています.まず1950年に産下された卵から成長した幼虫(1950年群)が,1951年の8月ごろ終齢になりました.この終齢幼虫は休眠に入り,夏,秋,冬と年を越して,翌年(1952年)5月17日以降15日間程度で同調的に羽化しました.一方で1951年に産下された卵から成長した幼虫(1951年群)の一部が春に終齢になって,そのまま発育遅延を起こさず,1950年群の羽化より少し遅れてだらだらと羽化しました.
そこで,彼は,日長の増減を幼虫は感じ取っていて,それに反応しているのではないかと考えました.そして詳細な実験計画を立て,この変化する日長の効果について検証しました.すると,長日から減少しつつある日長は休眠を誘導し,短日から増加しつつある日長は休眠を誘導しないという結果を得ました (Corbet,1956).ただ,温度条件が厳密にコントロールされていないため,さらなる検証が必要だとも言っています.
以上から,幼虫休眠は,原則的に日長によって誘導される外因性休眠といえそうです.しかし,「昆虫の光周性」を著したア・エス・ダニレフスキーは,「温度に対する生理的適応と日長に対する生理的適応とは,たがいに密接に関連し合っている.自然条件のもとでは,これら二つの要因の生態的役割を,べつべつに切り離して評価することは容易ではない.(日高・正木訳,1966より引用)」と述べています.トンボでも,日長と温度の組み合わせが幼虫の成長に影響を及ぼすという報告はあります(例えば Ingram & Jenner, 1976)が,休眠に対する光周期と温度の統合的役割については,まだ十分には解明されているようには見えません.今後の研究発展を期待したいと思います.
Ⅳ.成虫の休眠
成虫の休眠は,秋季種と成虫越冬種について見られる現象と考えてよいでしょう.これらのトンボは非常に長い前生殖期を有している点で,何らかの発育遅延が生じていることは疑いありません.しかし,休眠が起きているとして,日長なり温度なり,いかなる環境要因がそれを誘導しているのか,あるいは消去しているのか,さらに遺伝的に決められた内因性のものなのか,といったそのしくみについては,ほとんど報告がありません.これは成虫の継続飼育が困難なことが一番大きな原因だろうと推察されます.
したがって,野外でこの時期の状態を調べることになり,そういった研究はいくつか報告があります.Uéda (1989) は,金沢で夏越ししているアオイトトンボの前生殖期個体群を使って,その成熟の進行(体色変化,生殖巣の発達など)を調べています.それによると,オス・メスとも,前生殖期間を通じて,体色は遅延なく成熟状態へと変化していくようです.しかし生殖巣の発達についてはオス・メスで違いがあり,メスは8月中旬に至るまで成熟卵が現れないのに対し,オスの精子形成には遅延が起きず7月上旬には成熟していると報告しています.
これに関連して,兵庫県南部で,アオイトトンボが6月11日に連結産卵行動をとっているのが観察されています(青木,1991).ここでは羽化は5月中旬に始まっています.産卵痕を確認できていないため,卵がどういう状態だったかについての情報がないので,本当の産卵かどうかは不明です.しかしこれは,少なくともオスが遅延なく成熟している証かも知れません.
上田(1979)は,この繁殖期開始の遅れをもたらす環境要因について,日長の影響を否定しています.そして繁殖活動が開始されるのがほとんどの調査地域で24℃を下回ったときであることから,高温が繁殖活動を抑制している可能性を指摘しています.しかし一方で,高温がこの繁殖活動遅延を引き起こす引き金になるとはいえないとも述べています.それは,羽化して20日間(非休眠個体群が成熟する期間)過ぎた時期に,まだそれほど高温にならない地域があり,もし高温が繁殖活動を抑制する引き金になっているなら,この地域ではその時期に繁殖活動が始まってもおかしくないことになる,という考えに基づいています.
もう一つ,アルジェリアで調べられたタイリクアカネ原名亜種 Sympetrum striolatum および他2種の夏越しを調査した研究があります.これによると,やはりオス・メスとも体色は順次成熟色になっていくようですが,メスは9月下旬になってから成熟した卵を持つようになっています (Smuraoui et.al., 1998;Figure 5).
こういった研究から,少なくともメスにおいては,生殖巣の発育遅延が起きることがあるのは確かなようで,この現象は休眠(生殖休眠)であるといえるでしょう.成虫越冬種についても,まずこういった成熟の季節進行をきちんと押さえる研究を待ちたいと思います.
図9.Anax imperator の羽化前の幼虫の発育の模式図 (Corbet, 1955 の記述をもとに描画).
全体の羽化は5月17日から7月5日にかけて行われる.
最初の羽化ピークは15日間ほどで終了し,これは1951年夏から休眠に入った個体である.
遅れた羽化ピークは,その春に終齢になって,遅延することなく発育し羽化した個体である.
前者の終齢幼虫は休眠が誘導されているが,後者は誘導されない.
終齢幼虫には休眠するものとしないものがあるというこの結果から,この休眠が外因性休眠であることが分かります.そして,この休眠が日長によって引き起こされているならば,次のような疑問が生じると,述べています.夏至前の4-5月に終齢になった非休眠の幼虫が経験している日長は,終齢で夏以降休眠した幼虫も8-9月にに経験しているはずだから,この休眠誘導では日長の絶対長は重要ではない (Corbet, 1956).全体の羽化は5月17日から7月5日にかけて行われる.
最初の羽化ピークは15日間ほどで終了し,これは1951年夏から休眠に入った個体である.
遅れた羽化ピークは,その春に終齢になって,遅延することなく発育し羽化した個体である.
前者の終齢幼虫は休眠が誘導されているが,後者は誘導されない.
そこで,彼は,日長の増減を幼虫は感じ取っていて,それに反応しているのではないかと考えました.そして詳細な実験計画を立て,この変化する日長の効果について検証しました.すると,長日から減少しつつある日長は休眠を誘導し,短日から増加しつつある日長は休眠を誘導しないという結果を得ました (Corbet,1956).ただ,温度条件が厳密にコントロールされていないため,さらなる検証が必要だとも言っています.
以上から,幼虫休眠は,原則的に日長によって誘導される外因性休眠といえそうです.しかし,「昆虫の光周性」を著したア・エス・ダニレフスキーは,「温度に対する生理的適応と日長に対する生理的適応とは,たがいに密接に関連し合っている.自然条件のもとでは,これら二つの要因の生態的役割を,べつべつに切り離して評価することは容易ではない.(日高・正木訳,1966より引用)」と述べています.トンボでも,日長と温度の組み合わせが幼虫の成長に影響を及ぼすという報告はあります(例えば Ingram & Jenner, 1976)が,休眠に対する光周期と温度の統合的役割については,まだ十分には解明されているようには見えません.今後の研究発展を期待したいと思います.
Ⅳ.成虫の休眠
成虫の休眠は,秋季種と成虫越冬種について見られる現象と考えてよいでしょう.これらのトンボは非常に長い前生殖期を有している点で,何らかの発育遅延が生じていることは疑いありません.しかし,休眠が起きているとして,日長なり温度なり,いかなる環境要因がそれを誘導しているのか,あるいは消去しているのか,さらに遺伝的に決められた内因性のものなのか,といったそのしくみについては,ほとんど報告がありません.これは成虫の継続飼育が困難なことが一番大きな原因だろうと推察されます.
したがって,野外でこの時期の状態を調べることになり,そういった研究はいくつか報告があります.Uéda (1989) は,金沢で夏越ししているアオイトトンボの前生殖期個体群を使って,その成熟の進行(体色変化,生殖巣の発達など)を調べています.それによると,オス・メスとも,前生殖期間を通じて,体色は遅延なく成熟状態へと変化していくようです.しかし生殖巣の発達についてはオス・メスで違いがあり,メスは8月中旬に至るまで成熟卵が現れないのに対し,オスの精子形成には遅延が起きず7月上旬には成熟していると報告しています.
これに関連して,兵庫県南部で,アオイトトンボが6月11日に連結産卵行動をとっているのが観察されています(青木,1991).ここでは羽化は5月中旬に始まっています.産卵痕を確認できていないため,卵がどういう状態だったかについての情報がないので,本当の産卵かどうかは不明です.しかしこれは,少なくともオスが遅延なく成熟している証かも知れません.
上田(1979)は,この繁殖期開始の遅れをもたらす環境要因について,日長の影響を否定しています.そして繁殖活動が開始されるのがほとんどの調査地域で24℃を下回ったときであることから,高温が繁殖活動を抑制している可能性を指摘しています.しかし一方で,高温がこの繁殖活動遅延を引き起こす引き金になるとはいえないとも述べています.それは,羽化して20日間(非休眠個体群が成熟する期間)過ぎた時期に,まだそれほど高温にならない地域があり,もし高温が繁殖活動を抑制する引き金になっているなら,この地域ではその時期に繁殖活動が始まってもおかしくないことになる,という考えに基づいています.
もう一つ,アルジェリアで調べられたタイリクアカネ原名亜種 Sympetrum striolatum および他2種の夏越しを調査した研究があります.これによると,やはりオス・メスとも体色は順次成熟色になっていくようですが,メスは9月下旬になってから成熟した卵を持つようになっています (Smuraoui et.al., 1998;Figure 5).
こういった研究から,少なくともメスにおいては,生殖巣の発育遅延が起きることがあるのは確かなようで,この現象は休眠(生殖休眠)であるといえるでしょう.成虫越冬種についても,まずこういった成熟の季節進行をきちんと押さえる研究を待ちたいと思います.
トンボの生態学 7.生活史の季節的制御
同時出生集団の分割,発育の促進
生活史の季節的制御に重要な役割をしている,休眠の様相についてお話ししました.これ以外にも,生活史を特徴づける幼虫発育上の出来事があります.これが,「同時出生集団の分割」と「発育の促進」です.
同時出生集団の分割 cohort splitting とは,例えば通常二年一化するトンボで,ある年に孵化した幼虫が,2年で羽化するグループと1年で羽化するグループに分かれるような現象をいいます.具体例としては,図9で示した Anax imperator の1951年群の一部が分割して,他のものより1年早く羽化しています.つまり通常二年一化のものの一部から,一年一化のものが生じているということになります.
生活史の話からは少しそれますが,この現象にはとても重要な意味があります.例えば三年一化のトンボがいるとしましょう.このトンボが,厳格に三年一化が守られるような生活史が営まれているとすると,同一地域に,生殖隔離された(遺伝的に交流のない)3つの個体群が存在することになります.しかし,同時出生集団の分割が存在することで,これら各年齢群間の遺伝的交流を実現することが可能になります.図9でいうと,1952年の繁殖成虫の中には,1950年生まれのものとと1951年生まれのものが混じっているということです.
もう一つの発育の促進というのは,休眠が発育を遅延させて生活史をコントロールするのに対し,発育を促進させて生活史をコントロールするしくみのことです.これはまだあまり研究課題に上がったことがない現象です.
さて,同時出生集団の分割について考えるには,トンボ幼虫の成長のあり方に注意を向ける必要があります.幼虫は脱皮しながら大きくなっていきますが,一つは,孵化してから羽化までの齢数にはばらつきがあるということです.1頭のメスから産下された卵を飼育した研究で,例えば Aeshna cyanea では11齢,12齢,13齢,14齢で終齢になったものが,それぞれ6%,81%,11%,2%であったという記録があります(*Schaller, 1960).
二つ目は,脱皮の際のサイズの増加比(成長比)です.トンボ幼虫の頭幅長や下唇前基節長などは,通常アロメトリックな成長(等比数列的なサイズ増加)をすることが知られています.その成長比は多くのトンボで1.2から1.3の範囲にあります(例えば Corbet, 2002;Figure 5).したがって,例えば脱皮ごとに頭幅長を測定したものを片対数方眼紙にグラフ化すれば直線になります.そしてこの傾きが成長比(の対数をとったもの)を表します(図10参照).
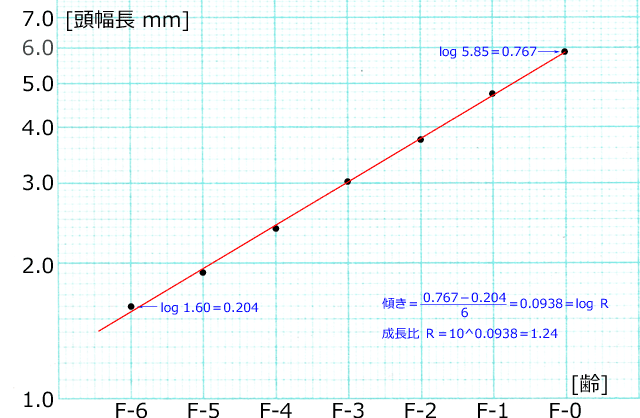
図10.サラサヤンマ Sarasaeschna pryeri 頭幅長のアロメトリックな成長.
縦軸は頭幅長平均値の対数,横軸は齢を表す.
各齢と頭幅長の対数は正比例する.つまり脱皮ごとに一定の比率で増加していることが分かる.
脱皮ごとの成長比は,だいたい1.25前後になっている.
元データは「幼虫がみつからない!? サラサヤンマの生態」のページ参照.
この成長比は,様々な要因,例えば,温度,光周期,餌の状態などが考えられますが,これらによって微妙に変化します.その結果,成長が進むにつれて,同じ齢でも,サイズにばらつきが出てくることになります.
同時出生集団の分割は,羽化前年の秋から羽化の直前の春にかけてみられることが多いようです.この分割の境界は,幼虫のサイズによって決まってくることが報告されています.つまり,羽化が近くなったある時点で,あるサイズより大きなものだけが羽化に向けて発育が促進され,そのサイズより小さなものは発育が遅延され羽化が一年先延ばしにされる,ということになります.Norling (1984) は,この境目になるサイズを冬季臨界サイズ winter critical size と名付けています.一般にこれはサイズ分布の極小値で見つけられるとも述べています(図11参照).
この様子をキイロサナエで見てみましょう.図11は,羽化前年の晩春から冬に至るまでの終齢(F-0)と亜終齢(F-1)のサイズ分布を示しています.同一年齢群のほとんどは,羽化2年前の冬を,F-2齢とF-3齢で過ごしています.F-2齢で越冬したものは晩春から初夏にかけてF-1齢へと脱皮します(図11の青色).F-3齢で越冬したものは,春にF-2齢となり,夏頃にF-1齢へと脱皮します(図11の赤色).後者は前者よりサイズが小さくなっていますが,これは成長が後れているものほど,その成長比がわずかに小さいためです(青木,1994).そしてこのF-1齢集団の大部分が秋に終齢(F-0)へと脱皮します.したがって,これらは冬のデータには表れていません.一方F-3齢で越冬した群由来の小さなF-1齢の一部は,終齢にならずF-1齢のまま越冬に入ります.このF-1齢はその後もう一度脱皮しますが,終齢にはならず,過剰脱皮したF-1齢になり,羽化が1年先送りされます.その過剰脱皮したF-1齢幼虫は,晩春から初夏のデータ,および夏のデータでも見いだすことができています(以上 Aoki, 1999).
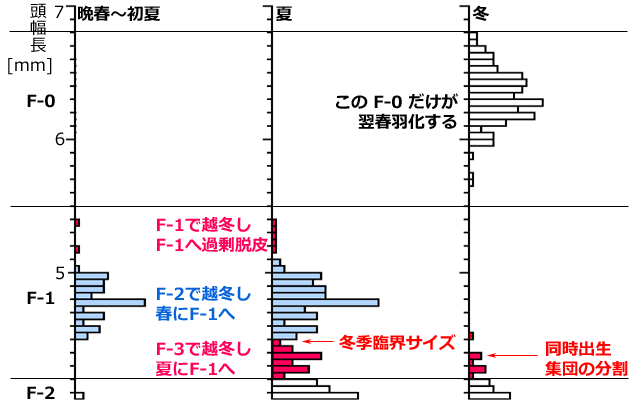
図11.キイロサナエの同時出生集団の分割と冬季臨界サイズ (Aoki, 1999を改変).
成長の様子がよく分かるように,3年間のデータを重ねて表示している.
晩春から初夏にかけてF-2齢で越冬した一群がF-1齢に脱皮(青色).
夏にはF-3齢で越冬した同じ年齢群がF-2齢を経てF-1齢に脱皮(赤色).
冬季臨界サイズは頭幅長で4.5mm前後にあると考えられる.
冬季臨界サイズ以上のF-1齢幼虫は秋に終齢へと脱皮し翌年春に羽化する(冬に見られない).
冬季臨界サイズ以下のF-1齢幼虫はそのまま越冬し,過剰脱皮してもう一年夏を過ごす.
キイロサナエは通常15齢が終齢だが,過剰脱皮した群は16齢が終齢となる.
以上からお分かりのように,羽化前年の夏から秋にかけての幼虫サイズが,翌年の羽化に進むか,もう一年過ごすかの分岐点を与えているということになるわけです.これが幼虫の齢数ではなくサイズで決められているという点が非常に面白いと感じます.キイロサナエの場合は,成長の後れている幼虫ほど成長比が小さくなる傾向が見いだされており(青木,1994),それらの幼虫に,一年長い生活史を営むことが生活史戦略として組み込まれているように思えます.また成長の後れた幼虫が羽化を1年先延ばしされることによって,羽化の終わる時期が設定され,それによって同調性がより高まるという点にも注目すべきでしょう.
次に「成長の促進」についてお話ししましょう.これは私も,Norling (2018) の論文で知ったことで,あまり考えたことがなかった状況です.この現象は,スェーデンの北緯55.75度のアオイトトンボ個体群を使って調べられました.南西日本では,アオイトトンボは夏に生殖休眠を行いますが,この地域ではそれは行われず,20日ほどの前生殖期のあと,繁殖活動を開始します.羽化は7月頃にピークを迎え,その後前生殖期間を経て産卵が始まります(図5参照.図5は北緯53.01度).緯度が高いこともあって冬の到来も早く,ここの個体群は繁殖活動にあてられる時間的余裕が少なくなるような状況に置かれています.したがって,幼虫の成長をできるだけ早く終え繁殖活動にあてる時間を稼ぐことは,適応的であると考えられます.
Norling (2018) は,日長をシフトさせる実験を行って,長日から短日へのシフトが幼虫の発育を促進することを見いだしました.野外で幼虫の成長する季節は夏至をはさんでその前後です.そこで彼は,同腹の卵から孵化した幼虫について,6齢を境に4つの組み合わせで日長をシフトさせました.明暗周期は,LD19.5:4.5(夏至の日長,以下19と表記)とLD16:8(5月・8月上旬の日長,以下16と表記)を用意し,シフト処理は16-16,16-19,19-19,19-16の4つです.するとこのうち「19-16」の実験群だけ羽化までの日数が有意に短くなったのです.
彼は,16-16という実験群の羽化までの日数が特に短くならなかったことから,LD16:8という短日だけが成長を加速したのではなく,LD19.5:4.5を最初の6齢で経験し,その後でLD16:8を経験したことが成長を促進させたと考えました.つまり幼虫は,夏至を経験し,だんだんと日が短くなっていくときに成長が促進され,羽化が早められる.その結果繁殖活動にあてられる時間を少しでも長くすることができる,という解釈になります.
さらに,19-19や19-16の実験群では,終齢への脱皮回数が減少する傾向もデータとして示しており,長日または減少する日長が,脱皮回数を減らす可能性を示唆しています.ただ彼も述べていますが,まだ実験の個体数が少なく,さらなる検証が必要だということです.
私は,幼虫の脱皮回数が,夏至の長日あるいは夏至を経験した後短くなりつつある日長によって減少する可能性に,とても興味を覚えました.一年二化するトンボの,二化目の個体がやや小さくなることについて(写真6),同様のメカニズムを考えることによって,これを説明できるのではないかと思ったからです.こういった仮説のもとに二化性トンボの生活史の研究をすると,面白い結果が出るかもしれません.
同時出生集団の分割 cohort splitting とは,例えば通常二年一化するトンボで,ある年に孵化した幼虫が,2年で羽化するグループと1年で羽化するグループに分かれるような現象をいいます.具体例としては,図9で示した Anax imperator の1951年群の一部が分割して,他のものより1年早く羽化しています.つまり通常二年一化のものの一部から,一年一化のものが生じているということになります.
生活史の話からは少しそれますが,この現象にはとても重要な意味があります.例えば三年一化のトンボがいるとしましょう.このトンボが,厳格に三年一化が守られるような生活史が営まれているとすると,同一地域に,生殖隔離された(遺伝的に交流のない)3つの個体群が存在することになります.しかし,同時出生集団の分割が存在することで,これら各年齢群間の遺伝的交流を実現することが可能になります.図9でいうと,1952年の繁殖成虫の中には,1950年生まれのものとと1951年生まれのものが混じっているということです.
もう一つの発育の促進というのは,休眠が発育を遅延させて生活史をコントロールするのに対し,発育を促進させて生活史をコントロールするしくみのことです.これはまだあまり研究課題に上がったことがない現象です.
さて,同時出生集団の分割について考えるには,トンボ幼虫の成長のあり方に注意を向ける必要があります.幼虫は脱皮しながら大きくなっていきますが,一つは,孵化してから羽化までの齢数にはばらつきがあるということです.1頭のメスから産下された卵を飼育した研究で,例えば Aeshna cyanea では11齢,12齢,13齢,14齢で終齢になったものが,それぞれ6%,81%,11%,2%であったという記録があります(*Schaller, 1960).
二つ目は,脱皮の際のサイズの増加比(成長比)です.トンボ幼虫の頭幅長や下唇前基節長などは,通常アロメトリックな成長(等比数列的なサイズ増加)をすることが知られています.その成長比は多くのトンボで1.2から1.3の範囲にあります(例えば Corbet, 2002;Figure 5).したがって,例えば脱皮ごとに頭幅長を測定したものを片対数方眼紙にグラフ化すれば直線になります.そしてこの傾きが成長比(の対数をとったもの)を表します(図10参照).
図10.サラサヤンマ Sarasaeschna pryeri 頭幅長のアロメトリックな成長.
縦軸は頭幅長平均値の対数,横軸は齢を表す.
各齢と頭幅長の対数は正比例する.つまり脱皮ごとに一定の比率で増加していることが分かる.
脱皮ごとの成長比は,だいたい1.25前後になっている.
元データは「幼虫がみつからない!? サラサヤンマの生態」のページ参照.
同時出生集団の分割は,羽化前年の秋から羽化の直前の春にかけてみられることが多いようです.この分割の境界は,幼虫のサイズによって決まってくることが報告されています.つまり,羽化が近くなったある時点で,あるサイズより大きなものだけが羽化に向けて発育が促進され,そのサイズより小さなものは発育が遅延され羽化が一年先延ばしにされる,ということになります.Norling (1984) は,この境目になるサイズを冬季臨界サイズ winter critical size と名付けています.一般にこれはサイズ分布の極小値で見つけられるとも述べています(図11参照).
この様子をキイロサナエで見てみましょう.図11は,羽化前年の晩春から冬に至るまでの終齢(F-0)と亜終齢(F-1)のサイズ分布を示しています.同一年齢群のほとんどは,羽化2年前の冬を,F-2齢とF-3齢で過ごしています.F-2齢で越冬したものは晩春から初夏にかけてF-1齢へと脱皮します(図11の青色).F-3齢で越冬したものは,春にF-2齢となり,夏頃にF-1齢へと脱皮します(図11の赤色).後者は前者よりサイズが小さくなっていますが,これは成長が後れているものほど,その成長比がわずかに小さいためです(青木,1994).そしてこのF-1齢集団の大部分が秋に終齢(F-0)へと脱皮します.したがって,これらは冬のデータには表れていません.一方F-3齢で越冬した群由来の小さなF-1齢の一部は,終齢にならずF-1齢のまま越冬に入ります.このF-1齢はその後もう一度脱皮しますが,終齢にはならず,過剰脱皮したF-1齢になり,羽化が1年先送りされます.その過剰脱皮したF-1齢幼虫は,晩春から初夏のデータ,および夏のデータでも見いだすことができています(以上 Aoki, 1999).
図11.キイロサナエの同時出生集団の分割と冬季臨界サイズ (Aoki, 1999を改変).
成長の様子がよく分かるように,3年間のデータを重ねて表示している.
晩春から初夏にかけてF-2齢で越冬した一群がF-1齢に脱皮(青色).
夏にはF-3齢で越冬した同じ年齢群がF-2齢を経てF-1齢に脱皮(赤色).
冬季臨界サイズは頭幅長で4.5mm前後にあると考えられる.
冬季臨界サイズ以上のF-1齢幼虫は秋に終齢へと脱皮し翌年春に羽化する(冬に見られない).
冬季臨界サイズ以下のF-1齢幼虫はそのまま越冬し,過剰脱皮してもう一年夏を過ごす.
キイロサナエは通常15齢が終齢だが,過剰脱皮した群は16齢が終齢となる.
次に「成長の促進」についてお話ししましょう.これは私も,Norling (2018) の論文で知ったことで,あまり考えたことがなかった状況です.この現象は,スェーデンの北緯55.75度のアオイトトンボ個体群を使って調べられました.南西日本では,アオイトトンボは夏に生殖休眠を行いますが,この地域ではそれは行われず,20日ほどの前生殖期のあと,繁殖活動を開始します.羽化は7月頃にピークを迎え,その後前生殖期間を経て産卵が始まります(図5参照.図5は北緯53.01度).緯度が高いこともあって冬の到来も早く,ここの個体群は繁殖活動にあてられる時間的余裕が少なくなるような状況に置かれています.したがって,幼虫の成長をできるだけ早く終え繁殖活動にあてる時間を稼ぐことは,適応的であると考えられます.
Norling (2018) は,日長をシフトさせる実験を行って,長日から短日へのシフトが幼虫の発育を促進することを見いだしました.野外で幼虫の成長する季節は夏至をはさんでその前後です.そこで彼は,同腹の卵から孵化した幼虫について,6齢を境に4つの組み合わせで日長をシフトさせました.明暗周期は,LD19.5:4.5(夏至の日長,以下19と表記)とLD16:8(5月・8月上旬の日長,以下16と表記)を用意し,シフト処理は16-16,16-19,19-19,19-16の4つです.するとこのうち「19-16」の実験群だけ羽化までの日数が有意に短くなったのです.
彼は,16-16という実験群の羽化までの日数が特に短くならなかったことから,LD16:8という短日だけが成長を加速したのではなく,LD19.5:4.5を最初の6齢で経験し,その後でLD16:8を経験したことが成長を促進させたと考えました.つまり幼虫は,夏至を経験し,だんだんと日が短くなっていくときに成長が促進され,羽化が早められる.その結果繁殖活動にあてられる時間を少しでも長くすることができる,という解釈になります.
さらに,19-19や19-16の実験群では,終齢への脱皮回数が減少する傾向もデータとして示しており,長日または減少する日長が,脱皮回数を減らす可能性を示唆しています.ただ彼も述べていますが,まだ実験の個体数が少なく,さらなる検証が必要だということです.
私は,幼虫の脱皮回数が,夏至の長日あるいは夏至を経験した後短くなりつつある日長によって減少する可能性に,とても興味を覚えました.一年二化するトンボの,二化目の個体がやや小さくなることについて(写真6),同様のメカニズムを考えることによって,これを説明できるのではないかと思ったからです.こういった仮説のもとに二化性トンボの生活史の研究をすると,面白い結果が出るかもしれません.
トンボの生態学 7.生活史の季節的制御
トンボの生活史の進化的考察
昆虫における生活史とその季節的制御の研究には,主に一年一化または一年多化の生活史を持つものが利用されています.トンボのように数年にわたる生活史を持つものを材料にした研究は多くありません.また草食性昆虫の場合,食草の季節変動との関連でコントロールがなされていることがあり,特定の餌にこだわらない肉食性のトンボとはかなり趣が異なる場合があります.
では,トンボはなぜ季節的制御がなされなければならないのでしょうか.その一つは,系統的(遺伝的)な制約があることだと考えられます.
トンボは熱帯で進化発展した昆虫であるとされています.長い乾季の存在するような地域では,トンボは乾季をどう乗り越えるかという課題に直面します.幼虫で乾季を乗り越えることは不可能ですから,そこでは卵または成虫で乗り越えるという生活史が進化した可能性があります.これがおそらく休眠性をともなう卵越冬や成虫越冬の起源でしょう.他方,一年を通じて降水量の多い湿潤な地域では,こういった進化は起きないだろうと思われます.
しかし,トンボが高緯度地方へ分布を拡大していくと,大きな問題になるのが冬の存在です.耐寒性のあるステージをどこに持ってくるかが,進化の鍵になります.現在の姿を見ればその結果は明らかで,ある種は成虫,ある種は卵,そして一番多くの種が採用しているのが幼虫です.
このうち卵越冬する秋季種については,雨季・乾季が存在する熱帯で,乾季を卵で乗り切るという生活史の枠組みが,温帯地方での卵越冬の前適応として存在していたと考えられます.また短い雨季の間に生じるような一時的な水たまりに生育するトンボの幼虫は,一般に発育の温度係数が高く,高い温度のもとで急速に成長する性質を持っているとされていますが,これは秋季種幼虫の速い成長と符合します.つまり,乾季が冬に,雨季が梅雨に対応して,生活史の枠組みが受け継がれているいると考えられるわけです(Corbet, 1962).この,冬を卵で越すという枠組みはなかなか変更が難しいようで,すでに述べたように,南西日本のアオイトトンボは成虫で生殖休眠まで行って,卵越冬の枠組みを維持しようとしています.
成虫越冬種も,同様に,乾期を成虫で過ごすという前適応があったのではないかと思われます.
では幼虫越冬種はどうでしょう.これについては,Norling (1984a) の興味深い総説があります.「北方に分布を広げているトンボの生活史の形態」というタイトルで,まさに,今考えようとしている内容そのものです.これを私なりに解釈してお話ししてみようと思います.以下は多くが Norling (1984a) のアイデアをもとにしていますが,私の考えも含まれています.
図12は,トンボの生活史において,耐寒性があるステージとないステージを,単純化して示したものです.卵に休眠性がなく幼虫越冬するトンボは,一般に卵,若齢幼虫,羽化前のアポライシス(内部で幼虫のクチクラから成体が分離していくこと)が始まった終齢幼虫,および成虫に耐寒性がないと考えられています(Corbet, 1999;Norling, 1984).またトンボの生活史の完了に必要な時間は種によって異なっているはずですが,非休眠性の成虫の寿命や,非休眠の卵の発生時間には大きな差はないと考えられるので,もっとも差が出るのは,幼虫の,それも耐寒性のある幼虫の継続期間であるといってよいでしょう.
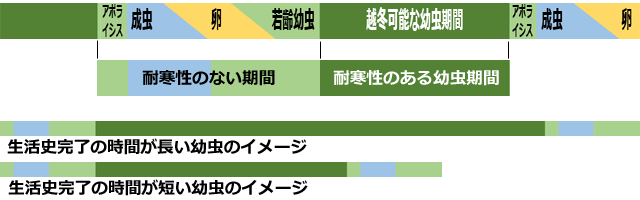
図12.幼虫で越冬するトンボの生活史と耐寒性の有無(単純化した模式図).
アポライシスを始めた終齢幼虫,成虫,卵,若齢幼虫には耐寒性がないとされている.
アポライシスとは,終齢幼虫体内で成虫への変態を始めた状態をさす.
生活史完了時間はトンボの種によって異なるが,それは主に幼虫期間の違いによる.
緯度が高くなってくると,温度は季節によって変動し,生活史の進行速度もその温度変化に影響され,変動するでしょう.ここでは温度による効果を平均化して,冬は発育が停止し,それ以外の活動可能な期間は一定の速度で発育が進むとして,生活史と季節変動の対応関係をシミュレーションしてみましょう.
さて,トンボが高緯度へ分布を広げていくときに最も重要なことは,耐寒性のある幼虫ステージを冬に合わせることです.その際,緯度による活動可能な期間の違いと,生活史完了に必要な時間の対応関係が,重要な意味を持ちます.例えば図13を見て下さい.これは,活動可能な期間と生活史の完了時間が一致している場合の例です.この場合,耐寒性のあるステージのどのタイミングで越冬に入っても,毎年同じタイミングで越冬に入ることができます.またこのタイミングには一定の範囲が存在しますので,幼虫は様々の齢期で越冬することになり,また成虫の出現時期にも幅が出てきます.これはちょうど夏季種の状態に似ています.
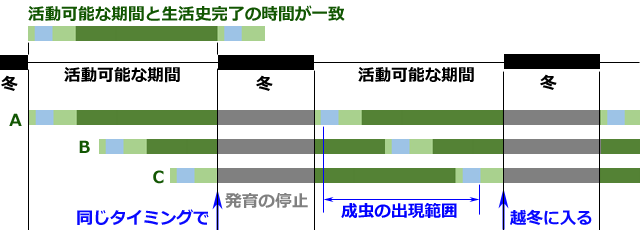
図13.活動可能期間と生活史完了時間が一致する場合のシミュレーション.
A.もっとも発育の進んだ幼虫で越冬する場合.
B.中間程度の発育状態の幼虫で越冬する場合.
C.もっとも発育の後れた幼虫で越冬する場合.
活動可能期間と生活史完了時間が一致しているので越冬に入るタイミングがずれない.
幼虫の越冬齢には幅ができ成虫の出現期間も長い.これは夏季種の生活史を実現している.
なお,寒さに感受性のあるステージが冬と重なった個体は淘汰されることになる.
図13の例では,特に幼虫に休眠性は必要なく,冬の間発育を休止するだけでよいことになります.そこで次に,生活史完了の時間が活動可能な期間より少し短い場合を考えてみましょう.この場合は,ある年に耐寒性のあるステージを冬に合わせても,次の年の発育が早く完了するために,越冬に入るタイミングにずれが生じてきます(図14).
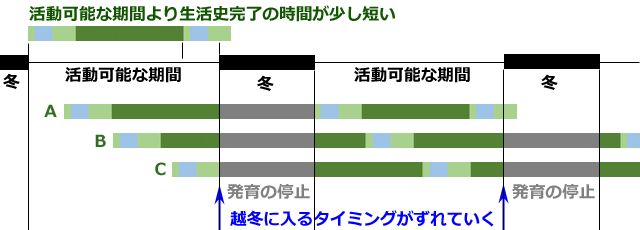
図14.活動可能期間より生活史完了時間が少し短い場合のシミュレーション.
A.もっとも発育の進んだ幼虫で越冬する場合.
B.中間程度の発育状態の幼虫で越冬する場合.
C.もっとも発育の後れた幼虫で越冬する場合.
活動可能期間より生活史完了時間が短いので越冬に入るタイミングがずれていく.
ずれが積み重なると,やがて図14Aのように耐寒性のないステージで越冬に入ることになり,死滅していまいます.したがってこの場合には,活動可能期間と生活史完了時間を一致させるためのしくみを獲得しなければなりません.これには二つの対処法が考えられます.一つは同じ温度に対する成長速度を落とす(発育の温度係数を下げる)ような進化です.そうすると幼虫の発育期間が長くなり生活史完了時間を活動可能期間に一致させることが可能になります.
発育の温度係数を変化させるこのやり方は,生活史完了時間が活動可能期間より少し長い場合には,逆に温度係数を高めることによって,これらを同調させることが可能になります.いずれにしてもその結果は図13と同じになり,夏季種のような生活史の実現につながります.現在夏季種がかなりの数いることを考えると,実際こういう方法を進化させているトンボはかなり存在するのではないかと思っています.
生活史完了時間の調節は,この温度係数の変化以外に,生息場所を選ぶことでも微調整できるかも知れません.平均気温が下がる場所では日当たりのよい水温が上がりやすい場所で生活するとか,またその逆もあり得るでしょう.ただし,活動可能期間と生活史完了時間のずれが大きくなると,これらの方法では,もはや調整ができなくなるだろうと想像されます.
もう一つの調節の方法は,休眠を間にはさんで,生活史完了の時間を延ばすことです.この場合は,活動可能期間と生活史完了の時間にかなり大きなずれがあっても,調整ができるはずです.理屈の上では,耐寒性のある幼虫期間のどこに休眠をはさみ込んでも,活動可能な期間と生活史完了時間の一致は実現できます.耐寒性のあるステージのすべてで休眠可能だとして,休眠期間が活動可能時間と生活史完了時間の差に等しい場合は,結局図13と同じになります.
現生種で,こういう事例が実際にあるかどうかは分かりませんので,これは理屈の上だけの話になります.そこで,現在よく見られる後期齢幼虫に外因性休眠がある場合をシミュレーションしてみましょう.
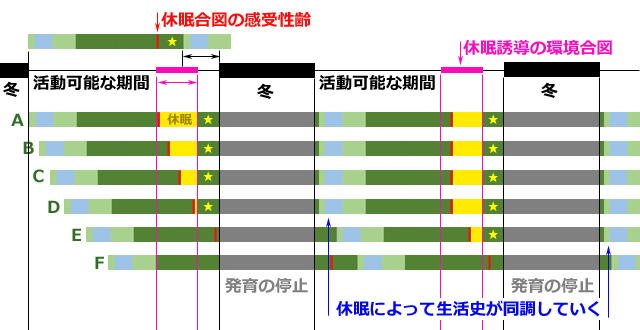
図15.活動可能期間より生活史完了時間が短い場合の休眠による調整シミュレーション.
A-F.すこしずつスタートをずらして休眠の効果を見た.
Fの2年目はEの1年目に近く,やがて生活史は同調していくことにある.
休眠が消去された場合は,休眠に入った時点から発育が再開されるとした(星印).
これはうまく生活史が調整された例を示している.
図15には,休眠による生活史の調整がうまくいった例を示しています.特に重要なのは,休眠期間の最大値を与える「休眠誘導の環境合図」の持続期間(図15桃色部)と,活動可能期間と生活史完了時間の差,それぞれの長さの関係です.前者が後者より短いと,調整はうまくいきません.
図15でもう一つ興味深い結果は,羽化の同調性が高まり,成虫の出現期間が春に同期してくる現象です.これは春季種の生活史を現出していることになります.このシミュレーションから考えると,春季種というのは,春に集中的に羽化することのメリットによって進化してきたというより,冬を耐寒性のある幼虫で越すために,後期齢に休眠を獲得・進化させた結果であるということになります.つまり春季種の登場は幼虫越冬のための進化の副産物ということになるのかも知れません.
もっと様々なシミュレーションが可能でしょうが,ここではこの一つだけにしておきます.おそらく現実の進化は,様々の休眠の枠組みがランダムに現れ,その中から図15のようにうまくいったものが選択されて,現在の生活史として残っているものと思われます.ここでは生活史完了時間が1年未満のものを扱いましたが,生活史完了時間が1年を超えるものについても,最終年またはその前年において調整されるということになるだけで,理屈は同様になります.
以上,熱帯で誕生したトンボが,まず熱帯で適応放散し,その生活史の枠組みを持ったまま高緯度へ分布拡大し,それぞれの場所で様々な生活史の「実験」を行い,自然選択が起きた結果,それぞれの地域の気候に適応していったと考えることができるでしょう.したがって,種内でも,緯度が違えば生活史やその季節的制御のあり方は大きくちがってくるでしょうし,逆に同所的に生活している異種間の生活史やその季節的制御のあり方が類似するということもあるでしょう.例えば現在日本で見られるような生活史の4つのパターンはそういう収斂の結果かも知れません.
Corbet (1999) には,unregulated development (制御されない発育)という語が出てきます.これは生活史のコントロールがされないで発育する幼虫のことです.例えば,熱帯のトンボであるウスバキトンボ幼虫は,高い温度係数のもとで,遅延なく発育を完了します.こういうトンボは他にもあるでしょう.しかし幼虫の発育には必ずばらつきが出てきて,シミュレーションのようなきちんとしたものにはなりません.そのため,冬の厳しい高緯度地方では,寒さに感受性のあるステージが冬と重ならないように,大なり小なり調整が必要だと考えられます.ただそれが非常に緩やかなものだと,春季種のように明瞭な制御がなされているものに比較すると,なかなか見いだすことが容易ではありません.まだまだトンボの季節適応には不明な点が多いのが実態です.
では,トンボはなぜ季節的制御がなされなければならないのでしょうか.その一つは,系統的(遺伝的)な制約があることだと考えられます.
トンボは熱帯で進化発展した昆虫であるとされています.長い乾季の存在するような地域では,トンボは乾季をどう乗り越えるかという課題に直面します.幼虫で乾季を乗り越えることは不可能ですから,そこでは卵または成虫で乗り越えるという生活史が進化した可能性があります.これがおそらく休眠性をともなう卵越冬や成虫越冬の起源でしょう.他方,一年を通じて降水量の多い湿潤な地域では,こういった進化は起きないだろうと思われます.
しかし,トンボが高緯度地方へ分布を拡大していくと,大きな問題になるのが冬の存在です.耐寒性のあるステージをどこに持ってくるかが,進化の鍵になります.現在の姿を見ればその結果は明らかで,ある種は成虫,ある種は卵,そして一番多くの種が採用しているのが幼虫です.
このうち卵越冬する秋季種については,雨季・乾季が存在する熱帯で,乾季を卵で乗り切るという生活史の枠組みが,温帯地方での卵越冬の前適応として存在していたと考えられます.また短い雨季の間に生じるような一時的な水たまりに生育するトンボの幼虫は,一般に発育の温度係数が高く,高い温度のもとで急速に成長する性質を持っているとされていますが,これは秋季種幼虫の速い成長と符合します.つまり,乾季が冬に,雨季が梅雨に対応して,生活史の枠組みが受け継がれているいると考えられるわけです(Corbet, 1962).この,冬を卵で越すという枠組みはなかなか変更が難しいようで,すでに述べたように,南西日本のアオイトトンボは成虫で生殖休眠まで行って,卵越冬の枠組みを維持しようとしています.
成虫越冬種も,同様に,乾期を成虫で過ごすという前適応があったのではないかと思われます.
では幼虫越冬種はどうでしょう.これについては,Norling (1984a) の興味深い総説があります.「北方に分布を広げているトンボの生活史の形態」というタイトルで,まさに,今考えようとしている内容そのものです.これを私なりに解釈してお話ししてみようと思います.以下は多くが Norling (1984a) のアイデアをもとにしていますが,私の考えも含まれています.
図12は,トンボの生活史において,耐寒性があるステージとないステージを,単純化して示したものです.卵に休眠性がなく幼虫越冬するトンボは,一般に卵,若齢幼虫,羽化前のアポライシス(内部で幼虫のクチクラから成体が分離していくこと)が始まった終齢幼虫,および成虫に耐寒性がないと考えられています(Corbet, 1999;Norling, 1984).またトンボの生活史の完了に必要な時間は種によって異なっているはずですが,非休眠性の成虫の寿命や,非休眠の卵の発生時間には大きな差はないと考えられるので,もっとも差が出るのは,幼虫の,それも耐寒性のある幼虫の継続期間であるといってよいでしょう.
図12.幼虫で越冬するトンボの生活史と耐寒性の有無(単純化した模式図).
アポライシスを始めた終齢幼虫,成虫,卵,若齢幼虫には耐寒性がないとされている.
アポライシスとは,終齢幼虫体内で成虫への変態を始めた状態をさす.
生活史完了時間はトンボの種によって異なるが,それは主に幼虫期間の違いによる.
さて,トンボが高緯度へ分布を広げていくときに最も重要なことは,耐寒性のある幼虫ステージを冬に合わせることです.その際,緯度による活動可能な期間の違いと,生活史完了に必要な時間の対応関係が,重要な意味を持ちます.例えば図13を見て下さい.これは,活動可能な期間と生活史の完了時間が一致している場合の例です.この場合,耐寒性のあるステージのどのタイミングで越冬に入っても,毎年同じタイミングで越冬に入ることができます.またこのタイミングには一定の範囲が存在しますので,幼虫は様々の齢期で越冬することになり,また成虫の出現時期にも幅が出てきます.これはちょうど夏季種の状態に似ています.
図13.活動可能期間と生活史完了時間が一致する場合のシミュレーション.
A.もっとも発育の進んだ幼虫で越冬する場合.
B.中間程度の発育状態の幼虫で越冬する場合.
C.もっとも発育の後れた幼虫で越冬する場合.
活動可能期間と生活史完了時間が一致しているので越冬に入るタイミングがずれない.
幼虫の越冬齢には幅ができ成虫の出現期間も長い.これは夏季種の生活史を実現している.
なお,寒さに感受性のあるステージが冬と重なった個体は淘汰されることになる.
図14.活動可能期間より生活史完了時間が少し短い場合のシミュレーション.
A.もっとも発育の進んだ幼虫で越冬する場合.
B.中間程度の発育状態の幼虫で越冬する場合.
C.もっとも発育の後れた幼虫で越冬する場合.
活動可能期間より生活史完了時間が短いので越冬に入るタイミングがずれていく.
発育の温度係数を変化させるこのやり方は,生活史完了時間が活動可能期間より少し長い場合には,逆に温度係数を高めることによって,これらを同調させることが可能になります.いずれにしてもその結果は図13と同じになり,夏季種のような生活史の実現につながります.現在夏季種がかなりの数いることを考えると,実際こういう方法を進化させているトンボはかなり存在するのではないかと思っています.
生活史完了時間の調節は,この温度係数の変化以外に,生息場所を選ぶことでも微調整できるかも知れません.平均気温が下がる場所では日当たりのよい水温が上がりやすい場所で生活するとか,またその逆もあり得るでしょう.ただし,活動可能期間と生活史完了時間のずれが大きくなると,これらの方法では,もはや調整ができなくなるだろうと想像されます.
もう一つの調節の方法は,休眠を間にはさんで,生活史完了の時間を延ばすことです.この場合は,活動可能期間と生活史完了の時間にかなり大きなずれがあっても,調整ができるはずです.理屈の上では,耐寒性のある幼虫期間のどこに休眠をはさみ込んでも,活動可能な期間と生活史完了時間の一致は実現できます.耐寒性のあるステージのすべてで休眠可能だとして,休眠期間が活動可能時間と生活史完了時間の差に等しい場合は,結局図13と同じになります.
現生種で,こういう事例が実際にあるかどうかは分かりませんので,これは理屈の上だけの話になります.そこで,現在よく見られる後期齢幼虫に外因性休眠がある場合をシミュレーションしてみましょう.
図15.活動可能期間より生活史完了時間が短い場合の休眠による調整シミュレーション.
A-F.すこしずつスタートをずらして休眠の効果を見た.
Fの2年目はEの1年目に近く,やがて生活史は同調していくことにある.
休眠が消去された場合は,休眠に入った時点から発育が再開されるとした(星印).
これはうまく生活史が調整された例を示している.
図15でもう一つ興味深い結果は,羽化の同調性が高まり,成虫の出現期間が春に同期してくる現象です.これは春季種の生活史を現出していることになります.このシミュレーションから考えると,春季種というのは,春に集中的に羽化することのメリットによって進化してきたというより,冬を耐寒性のある幼虫で越すために,後期齢に休眠を獲得・進化させた結果であるということになります.つまり春季種の登場は幼虫越冬のための進化の副産物ということになるのかも知れません.
もっと様々なシミュレーションが可能でしょうが,ここではこの一つだけにしておきます.おそらく現実の進化は,様々の休眠の枠組みがランダムに現れ,その中から図15のようにうまくいったものが選択されて,現在の生活史として残っているものと思われます.ここでは生活史完了時間が1年未満のものを扱いましたが,生活史完了時間が1年を超えるものについても,最終年またはその前年において調整されるということになるだけで,理屈は同様になります.
以上,熱帯で誕生したトンボが,まず熱帯で適応放散し,その生活史の枠組みを持ったまま高緯度へ分布拡大し,それぞれの場所で様々な生活史の「実験」を行い,自然選択が起きた結果,それぞれの地域の気候に適応していったと考えることができるでしょう.したがって,種内でも,緯度が違えば生活史やその季節的制御のあり方は大きくちがってくるでしょうし,逆に同所的に生活している異種間の生活史やその季節的制御のあり方が類似するということもあるでしょう.例えば現在日本で見られるような生活史の4つのパターンはそういう収斂の結果かも知れません.
Corbet (1999) には,unregulated development (制御されない発育)という語が出てきます.これは生活史のコントロールがされないで発育する幼虫のことです.例えば,熱帯のトンボであるウスバキトンボ幼虫は,高い温度係数のもとで,遅延なく発育を完了します.こういうトンボは他にもあるでしょう.しかし幼虫の発育には必ずばらつきが出てきて,シミュレーションのようなきちんとしたものにはなりません.そのため,冬の厳しい高緯度地方では,寒さに感受性のあるステージが冬と重ならないように,大なり小なり調整が必要だと考えられます.ただそれが非常に緩やかなものだと,春季種のように明瞭な制御がなされているものに比較すると,なかなか見いだすことが容易ではありません.まだまだトンボの季節適応には不明な点が多いのが実態です.
トンボの生態学 7.生活史の季節的制御
いくつかのトンボの生活史と季節的制御
では,この章の最後に,いくつかの種について具体的な生活史や,その季節的制御のあり方をまとめておきましょう.なお,参照した文献はすでに紹介しているので,ここでは新出文献を除いて,出典を割愛しています.
ではまずアオイトトンボについてまとめてみましょう.アオイトトンボは卵越冬の枠組みを維持しているトンボです.秋に産下された卵は内因性の休眠を行って越冬します.イギリスの研究では10℃15週間程度で休眠発育を終えます.兵庫県南部では5月から羽化を始め,生殖休眠に入ります.このとき,オスの体内では精子形成が遅延なく進んでいますが,メスの卵巣の成熟に遅延が発生し,金沢の個体群では8月中旬ころから成熟してきます.繁殖活動が開始される環境の合図は日長ではなく温度のようで,24℃を下回ることから始まります.あとは,繁殖能力が続く限り産卵が行われ,やがて成虫は,兵庫県南部では11月上旬までに消えていきます.
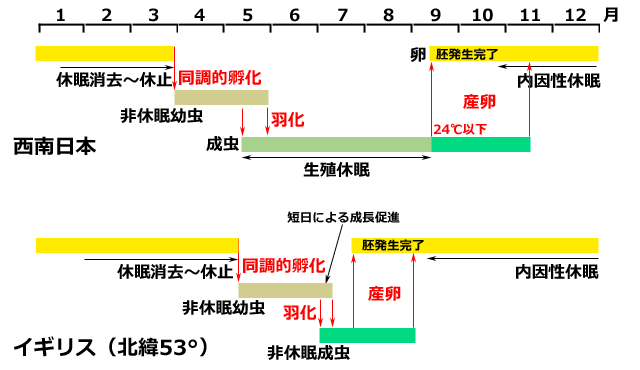
図16.アオイトトンボの生活史(上:南西日本の典型,下:北緯53度のイギリス)
アオイトトンボは卵越冬をする厳格な一年一化の生活史の枠組みを持っている.
卵は胚発生を完了した段階で内因性休眠に入る.
冬の間に休眠発育を終え,低温による休止段階に入って温度が孵化閾値を越えるのを待つ.
発育がそろっているので同調的に孵化し,幼虫は非休眠の温度依存的発生を行う.
羽化した成虫は,西南日本では生殖休眠を行うが,高緯度地方ではそれがない.
西南日本で生殖休眠が解けて繁殖活動を開始するのは,24℃を下回ったころである.
ではまずアオイトトンボについてまとめてみましょう.アオイトトンボは卵越冬の枠組みを維持しているトンボです.秋に産下された卵は内因性の休眠を行って越冬します.イギリスの研究では10℃15週間程度で休眠発育を終えます.兵庫県南部では5月から羽化を始め,生殖休眠に入ります.このとき,オスの体内では精子形成が遅延なく進んでいますが,メスの卵巣の成熟に遅延が発生し,金沢の個体群では8月中旬ころから成熟してきます.繁殖活動が開始される環境の合図は日長ではなく温度のようで,24℃を下回ることから始まります.あとは,繁殖能力が続く限り産卵が行われ,やがて成虫は,兵庫県南部では11月上旬までに消えていきます.
図16.アオイトトンボの生活史(上:南西日本の典型,下:北緯53度のイギリス)
アオイトトンボは卵越冬をする厳格な一年一化の生活史の枠組みを持っている.
卵は胚発生を完了した段階で内因性休眠に入る.
冬の間に休眠発育を終え,低温による休止段階に入って温度が孵化閾値を越えるのを待つ.
発育がそろっているので同調的に孵化し,幼虫は非休眠の温度依存的発生を行う.
羽化した成虫は,西南日本では生殖休眠を行うが,高緯度地方ではそれがない.
西南日本で生殖休眠が解けて繁殖活動を開始するのは,24℃を下回ったころである.

写真12.日本のエゾイトトンボ Coenagrion lanceolatum.Coenagrion hastulatum と似ている.
次に,夏季種に分類される,エゾイトトンボ属の Coenagrion hastulatum の生活史を紹介してみましょう(Norling, 1984).研究場所は北緯58.80度のスェーデンです.夏季種ではありますが,夏が短いこの高緯度の生息地では羽化は初夏に同調的に行われています.羽化群は,羽化直前の冬を終齢,F-1齢,F-2齢で過ごしています.このイトトンボは,羽化した年に,成長の速い幼虫と遅い幼虫に分かれ,若い齢期で同時出生集団の分割が起きます.その後成長の速い一群は主にF-1齢またはF-2齢で越冬し,翌春の長日で発育が促進され羽化します.この一群は一年一化の生活史になります.一方羽化の年の成長が遅かった一群は,翌年の夏の長日で発育が遅延され,終齢幼虫で越冬に入ります.そして翌春早々に羽化します.こちらは二年一化です.したがって,羽化する一群には異なる年齢群の個体が混じっていることになります.
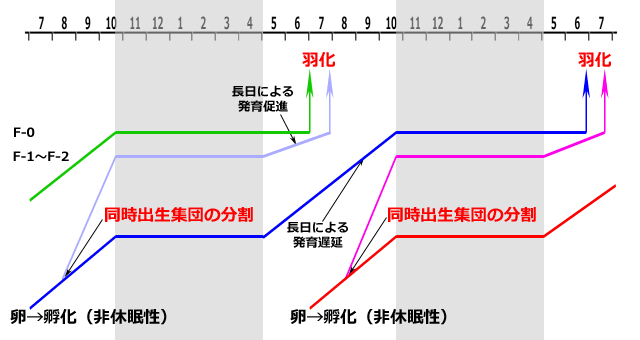
図17.Coenagrion hastulatum の生活史(スェーデン,北緯58.80度)(Norling, 1984より)
孵化した年のかなりの若齢段階で,同時出生群分割が始まる.
それによって,二年一化の群と一年一化の群に分かれる.
最後に,春季種のキイロサナエの生活史を紹介します.キイロサナエの卵には休眠性はありません.孵化した幼虫は,通常3回冬を越して,4年目の春に羽化します.1,2年目には目立った発育の遅延は見られず,3年目のF-1齢幼虫に,長日による発育遅延が見られます.これは秋の中間日長で消去され,一斉に終齢幼虫に脱皮します.このときに,同時出生群の分割が見られ,サイズの小さな幼虫は翌年過剰脱皮して一年羽化が先延ばしされます.秋に終齢へ一斉に脱皮することと,同時出生群分割によって成長の後れている幼虫の羽化が先延ばしされることで,羽化の同調性は非常に高くなります.
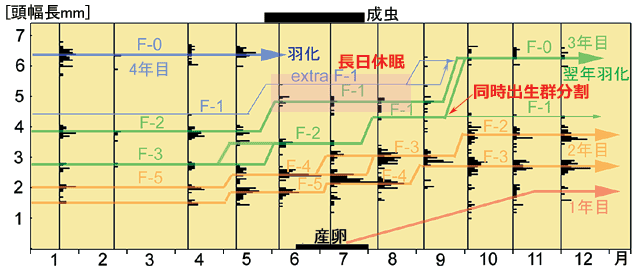
図17.キイロサナエ幼虫の発育(青木,1997より).
キイロサナエは,多くの幼虫が3回越冬し4年目の春に羽化する.
同じ年齢群のうち一部は同時出生集団の分割によって,4回越冬し5年目の春に羽化する.
6,7,8月には終齢幼虫が出現しない.これはF-1齢幼虫に長日休眠が生じているからである.
この長日休眠は9月頃に消去され,同調的に終齢幼虫になる.
2年目のF-4,F-3齢幼虫の頭幅長は,成長が後れているものほど小さくなっている.
生活史の季節的制御というと,生活史が厳格にコントロールされるという印象になります.Norling (1984a) には,北緯58度42分と北緯67度50分(北極圏!)におけるルリボシヤンマの生活史が紹介されていて,これによると,初年の卵越冬を含め,二年一化から五年一化までの生活史の変異があることが示されています.その年々の気象条件によって,またより北への分布拡大によって,生活史の枠組みが柔軟に変えられることが分かります.すべてのトンボがそうではないと思いますが,変更可能な可塑性と,最後の方できちんと羽化に向かって帳尻を合わせる厳密さ,この両者をうまく使って生活史を新しい生活場所に合わせながら分布を広げているのがトンボというものなのだと思います.
図17.Coenagrion hastulatum の生活史(スェーデン,北緯58.80度)(Norling, 1984より)
孵化した年のかなりの若齢段階で,同時出生群分割が始まる.
それによって,二年一化の群と一年一化の群に分かれる.
図17.キイロサナエ幼虫の発育(青木,1997より).
キイロサナエは,多くの幼虫が3回越冬し4年目の春に羽化する.
同じ年齢群のうち一部は同時出生集団の分割によって,4回越冬し5年目の春に羽化する.
6,7,8月には終齢幼虫が出現しない.これはF-1齢幼虫に長日休眠が生じているからである.
この長日休眠は9月頃に消去され,同調的に終齢幼虫になる.
2年目のF-4,F-3齢幼虫の頭幅長は,成長が後れているものほど小さくなっている.
参考文献
日高敏隆・正木進三訳,1966.「ア・エス・ダニレフスキー,1961.昆虫の光周性」.東京大学出版会.
Ando, H., 1962. The comparative embryology of Odonata with special reference to a relic dragonfly Epiophlebia superstes Selys. Jap. Soc. Prom. Sci., Tokyo.
安藤 裕,1996.トンボ目.安藤 裕・小林正幸監修 昆虫発生学[上].培風館.175-192.
安藤喜一,2004.イナゴ類の卵休眠と季節適応.休眠の昆虫学:4-15.,東海大学出版会.
青木典司,1991.アオイトトンボの6月の産卵行動.Gracile 45:18-19.
青木典司,1994.キイロサナエ幼虫の成長(第2報)-卵期間,幼虫期間,全齢数,羽化-.Tombo 37(1/4):31-36.
青木典司,1997.神戸のトンボ.神戸市スポーツ教育公社.
Aoki, T., 1999: Larval development, emergence and seasonal regulation in Asiagomphus pryeri (selys) (Odonata: Gomphidae). Hydrobiologia 394:179-192.
*Boehum, C., 1971. The influence of temperature upon embryonic diapause and seasonal regulation in Sympetrum vicinum (Hagen) (Odonata; Libellulidae). DrT, Univ. North Carolina, Chapel Hill. (in corbet, 1999)
Corbet, P.S., 1955. A critical response to changing length of day in an insect. Nature 175:338.
Corbet, P.S., 1956a. Environmrntal factors influencing the induction and termination of diapause in the emperor dragonfly Anax imperator Leach (Odonata: Aescnidae). The Journal of Experimental Biology 33(1):1-14.
Corbet, P.S., 1956b. The influence of temperature on diapause development in the dragonfly Lestes sponsa (Hansemann) (Odonata: Lestidae). Proc. R. ent. Soc. Lond. (A) 31:45-48.
Corbet, P.S., 1962. A Biology of Dragonflies. H. F. & G. Witherby ltd., GB.
Corbet, P.S., 1999. Dragonflies Behavior and Ecology of Odonata. Cornell Universitiy Press. New York.
Corbet, P.S., 2002. Stadia and growth ratios of Odonata: a review. International Journal of Odonatology 5(1):45-73.
Ingram, B.R. & C.F. Jenner, 1976. Life history of Enallagma hageni (Walsh) and E. aspersum (Hagen) (Zygoptera: Coenagrionidae). Odonatologica 5(4):331-345.
関西トンボ談話会,1984.近畿のトンボ.
近藤祥子,1994.数種のトンボの卵期・幼虫期の記録.Aeschna 29:15-20.
正木進三,1974.昆虫の生活史と進化.中公新書350.
Miller P.L., 1992. The effect of oxygen lack on egg hatching in an Indian dragonfly, Potamarcha congener. Physiol. Entomol. 17:68-72. (Abstruct).
Norling, U., 1976. Seasonal regulation in Leucorrhinia dubia (Vander Linden) (Anisoptera: Llibellulidae). Odonatologica 5:245-263.
Norling, U., 1984a. Life history patterns in the northern expansion of dragonflies. Adv. Odonatl. 2:127-156.
Norling, U., 1984b. The life cycle and larval photoperiodic respopnses of Coenagrion hastulatum (Charpentier) in two climatically different areas (Zygoptera: Coenagrionidae). Odonatologica 13(3):429-449.
Norling, U. & Sahlén G., 1997. Odonata, Dragonflies and Damselflies. Aquatic Insects of North Europe - A taxonomic Handbook. Volume 2:14-65. Apollo Books, Denmark.
尾園 暁・川島逸郎・二橋 亮,2012.ネイチャーガイド 日本のトンボ.文一総合出版,東京.
Samraoui, B., S. Bouzid, R. Boulahbal & P.S. Corbet, 1998. Postponed reproductive maturation in upland refuges maintain life-cycle continuity during the hot, dry seazon in Algelian dragonflies (Anisoptera). International Journal of Odonatology 1(2):118-135.
*Sawchyn W.W., and Church, C., 1973. The effects of temperature and photoperiod on diapause development in the eggs of four species of Lestes (Odonata; Zygoptera). Canadian Journal of Zoology, 51:1257-1265. (in Corbet, 1999)
*Sawchyn W.W., and Gillott, C., 1974. The lifehistories of three species of Lestes (Odonata; Zygoptera) in Saskatchewan. Camadian Entomologist, 106:1283-1293. (in Corbet, 1999)
*Schaller, F., 1960. Étude du développement postembryonnaire d'Aeshna cyanea. Mull. Ann. Sci. Nat. Zool. Biol. Anim. 2:751-868.(in Corbet,1999).
新村捷介・近藤祥子,1989.アカトンボの卵期及び卵の大きさについて.Gracile 42:1-5.
園部治之,1990.カイコの胚休眠-ホルモンと遺伝子による調節.昆虫生理学 156-178. 朝倉書店.
上田哲行,1979.アオイトトンボの生活史の起源.昆虫と自然 14(6):23-27.
Ubukata, H., 1980. Life History and Behaviour of a Corduliid Dragonfly Cordulia aenea amurensis SELYS. III. Aquatic Period, with Special References to Larval growth. Kontyu, Tokyo, 48(3):414-427.
Uéda, T., 1978. Geographic variation in the life cycle of Lestes sponsa. Tombo 21(1/4):27-34.
上田哲行,1990.トンボ類の"autumn species"の生活史 -その特性と変異-.個体群生態学会会報 46:62-67.
上田哲行,1993.山へ上るアキアカネ上らないアキアカネ.アキアカネの生活史における諸問題-1.インセクタリウム 30(9月号):4-11.
上田哲行,1996.アカトンボ類の多様な繁殖行動をどう理解するか.昆虫と自然 31(8):2-8.
吉田雅澄,2014.飼育による日本産トンボの卵・幼虫期.Aeschna (50):1-12.