トンボの生態学
4.卵と胚発生 目次
トンボの生態学 4.卵と胚発生
卵形成から胚発生完了までの概略
卵や胚発生には独特の用語がたくさん出てきます.1個の細胞である卵が細胞分裂し,さまざまな組織や器官を形成していきますので,発生が進み組織が分化していくと,それらに次々と新しい名称が付与されていきます.しかも同じ場所の名前が,新たに分化が進むと別の名称に変わったりします.成虫の組織や器官とその名称は,原則的に一対一に対応していますので,それらを覚えるだけで事足りますが,発生学においては,それに時間軸が加わって名称が変わったり,増えていくことになるわけです.
ここでは,トンボの発生学の詳細については専門書 (例えば,安藤 (1991);安藤・小林監修 (1996) など) に譲るとしてあまりに深入りすることはしないつもりですが,それでも高校生物の知識程度には見ていこうと考えています.そこで,まずはトンボの卵の成熟と胚発生に関する基本的順序を模式的に示して概念的にトンボの発生順序と基本的用語を理解していただこうと考えました.
なお,筆者は卵や胚発生に関する写真をほとんど持っていません.これに関しては
「近畿のトンボ雑記」という非常に素晴らしいWebサイトがありますので,それを参照してください.また
「神戸のトンボ」に,渡辺庸子氏にご提供いただいた素晴らしい写真がありますので,それも参照してください.
Ⅰ.卵とその成熟
卵は卵巣でつくられ成熟します.卵巣の中にある細胞のうち将来卵になる可能性を持った細胞を
卵原細胞と呼んでいます.この細胞の染色体数 (核相) は
2n で,同じ染色体を2本ずつ持っています.よくご存知の通り,1本は父親のオスから,もう1本は母親のメスから受け継いでいます.この卵原細胞は体細胞分裂を行って数を増やしています.
卵原細胞の中から,いよいよ卵になる運命を背負う
卵母細胞になるものが現れてきます.この卵母細胞は次の細胞分裂のとき,配偶子をつくるための細胞分裂,すなわち
減数分裂を行います.減数分裂は2回続けておき,最終的に4個の細胞ができます.精子の場合はこれらは4個の精子になっていきますが,卵の場合は,このうち1個だけが卵になり,他の3つは
極体と呼ばれる小さな細胞となって,やがて消失していきます.いずれにしても,減数分裂によって染色体数は半減し,核相は
n になります.
トンボの卵巣は
無栄養型と呼ばれるタイプに分類されます.これは卵に栄養を与える栄養細胞が分化しないタイプの卵巣です.代わりに
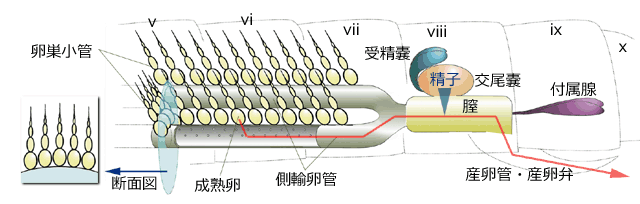
濾胞細胞が,卵殻などの卵の付属物を形成したり,卵に栄養を与えています.卵巣内で卵が形成される場所である
卵巣小管は欄外図1のような形状をしています.いちばん根元の柄部は
輸卵管 (側輸卵管) につながっており,柄部に隣接した卵がもっとも成熟度が高い卵です (欄外図1).この卵が成熟すると輸卵管に排出されます.これが
排卵です.するとその先に未熟な卵が次々に基部の方に移動します.
このように卵巣小管を移動しながら卵は成熟していきます.卵が成熟するこの部位を
卵黄巣と呼びます.ところで,卵の成熟とは何を意味するのでしょうか.それは,卵内に受精後の胚発生に必要となる,
卵黄やRNAなどの物質が蓄えられ卵内の適切な位置に配置されること,また卵を包む堅い卵殻などが形成されることを言います.このあたりの詳細はショウジョウバエを使って教科書に載るほどよく知られています.トンボに関してはほとんど研究されていませんが,濾胞細胞がこれらのはたらきをつかさどっていると考えられます.
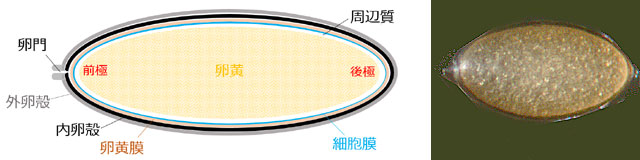
排卵される直前の成熟卵は,図2のような構造をしています.卵はそれ自体で一つの細胞ですので,卵全体を包む
細胞膜があります.そのすぐ外側に
卵黄膜があって,それを包むように堅い
卵殻があります.卵殻は
内卵殻と
外卵殻の2層からなっています.卵黄膜,内卵殻,外卵殻をまとめて
卵膜といいます.トンボの卵には透明なジェリー様物質が卵の外側に広がっているものがありますが,これは外卵殻が変化したものです (Corbet, 1999).昆虫の卵は
心黄卵といって卵の中心部に卵黄が分布しています.卵の
周辺質は細胞膜と卵黄の薄い隙間に卵黄を取り囲むように存在していて,顕微鏡ではほとんど見えません.
前極というのは,減数分裂のときに極体が放出される側で,通常精子が通り抜ける卵核の穴である
卵門が存在します.卵の核 (卵核) はこちら側の細胞質内に偏在しています.これは卵門を通り抜けて,細胞内に入り込んだ精子の核 (精核) と合体しやすいからでしょう.
卵門の数はトンボの種によって異なっており,例えば不均翅亜目・ムカシトンボ亜目においては,ムカシトンボで12~14個,ヤンマ科で5~10個,エゾトンボ科で2~4個,トンボ科で2個と,古い系統ほどその数が多くなっています (Becnel & Dunkle, 1990).
図2.トンボの卵の構造 (模式図)とキトンボの卵.
模式図はキトンボとは無関係である.周辺質とは細胞質のうち卵黄を除いた部分と考えてよい.卵殻は外卵殻と内卵殻からなり,その内側の卵黄膜と合わせて卵膜と呼ぶ.細胞膜は卵膜の内側にある.
卵の構造物は,植物内産卵・植物外産卵を行うものの間で若干の相違があります.一般に植物外産卵を行う種の卵は,卵黄膜が非常に分厚く内卵殻が薄くなっており,外卵殻はジェリー層に変化しています.植物内産卵を行う種はほぼ図2の通りで,卵黄膜はそれほど分厚くなく内卵殻は多層構造をしており,外卵殻には弾力性があるということです (Corbet, 1999).
排卵された成熟卵は側輸卵管を通り
膣に入ります.膣には
受精嚢や
交尾嚢といわれるオスの精子を蓄えている器官の口が開いており,この中の精子が,膣の通過時に卵に降りかけられ,卵門を通って卵内に進入し,卵黄膜に達します.卵黄膜に到達した精子はそれを溶かし細胞膜へと進んでいきますが,卵内に入るのはそのうちの一つだけです.この選ばれた精子は自身の細胞膜と卵の細胞膜を癒合させ,精核を細胞質内に注入します.精核は移動し卵核と合体し,この時点で
受精が完了します.
図3.トンボの卵巣の模式図.青木 (1997) を改変.
交尾嚢には過去に交尾したオスの精子が貯蔵されている.成熟卵が膣を通り抜けるとき,精子がふりかけられ,卵門から進入する.このとき卵は減数第一分裂の途中である.
膣を通った卵は,産卵管や産卵弁を通じて,しかるべき産卵基質に置かれます.先に減数分裂の話をしましたが,成熟卵は減数第一分裂の中期というステージで止まっており,精子が進入したころに減数第一分裂,さらに減数第二分裂を終えて,卵核が形成されるというタイミングになっています.また精子が進入した後,精核の形成にも多少の時間がかかりますので,実際の精核と卵核の合体は,卵が置かれた後しばらく時間が経ってからということになります.そして受精卵は発生を始めます.
Ⅱ.胚発生
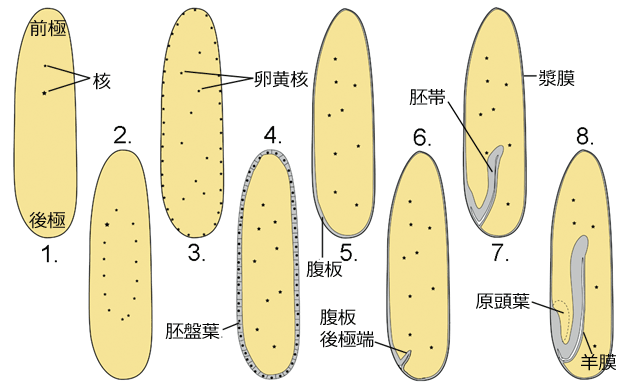
受精卵は細胞分裂 (
卵割) を始めます.みなさんが学校で習った細胞分裂は,核分裂が起きたあとすぐに細胞質分裂が起きて,細胞が二つ,四つ,八つになっていくものだったと思います.トンボなど昆虫の卵割は,まず核分裂だけが先行して何回も起きます.つまり卵は多核状態になります (図4-1, 2).ムカシトンボの卵では,第五卵割のときに多くの核が細胞表面に移動を始めます.そして第七卵割のころに核は細胞表面に到達し (図4-3),続く第八卵割で表面の核のまわりに細胞質分裂が生じ,薄い一層の細胞層が卵膜の内側に形成されます (図4-4).これを
胚盤葉と呼びます.卵黄内に残された核は卵黄細胞の核 (卵黄核) になります.このような卵割様式を
表割と呼びます.胚盤葉が形成されるあたりまで,核分裂はほぼ同調的に起きます (以上,安藤,1996).
図4.卵割と胚帯形成 (ムカシトンボ).青木 (1997) を改変.原図は Ando (1962) を参照.
さらに時間が経つと,胚盤葉の後極に近い部分に白く透き通った部分が現れてきます.これを
腹板と呼び,胚が形成され始めた場所になります (図4-5).腹板では盛んに細胞分裂が起き細胞数が増えていき,やがて腹板の後極端は卵黄の内部に陥入していきます (図4-6).この腹板の陥入部を
胚帯と呼びます (図4-7).陥入時に引っぱり込まれた漿膜の一部は
羊膜へと分化していきます(図4-8).なお胚帯の羊膜に近い面が胚の腹面になります.また原頭葉の部分が頭部になり,陥入部の胚帯先端が尾部になります.
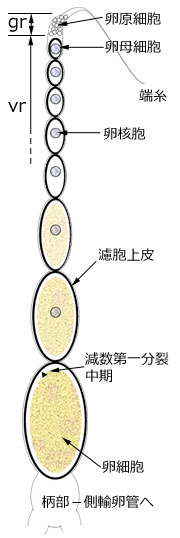
図1.卵巣小管の模式図.
濾胞上皮 (図の黒太線) は1層の濾胞細胞からなり,卵殻などの付属物の形成,栄養補給を行う.卵原細胞を含む部分 gr は形成細胞巣と呼ばれる.卵核胞は減数分裂前の核で卵が未熟な証拠となる.いちばん基部の成熟卵にはこれがなく,減数分裂が始まっている.柄部は成熟卵が側輸卵管に出る出口.端糸は卵巣小管が背壁に付着する部位.卵が成熟する部分 vr は卵黄巣と呼ばれる.
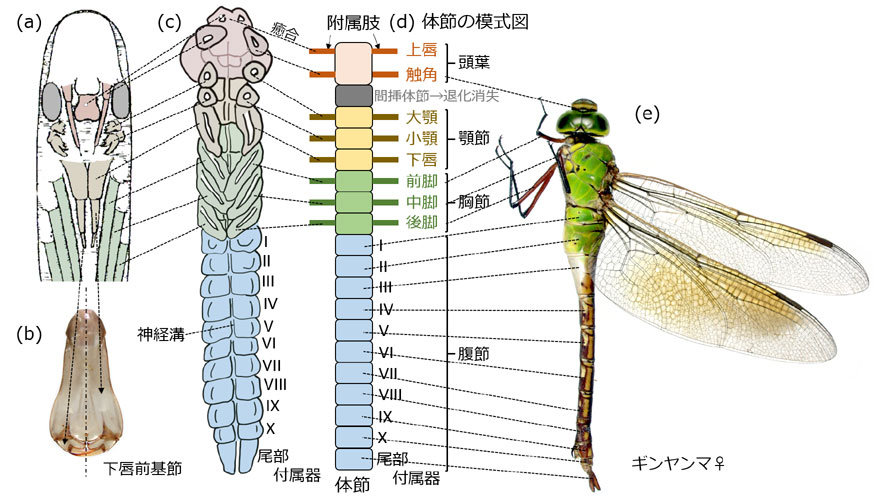
胚帯はどんどん伸長を続け,その間に胚帯には体節が形成され始めます.体節というのは進化した多細胞生物に共通の非常に重要な構造で,体軸方向の体をつくるための座標となるものです*1.つまり体節ごとに決まった器官が形成されて,正しい身体が形づくられていくのです.各体節には一対の付属肢と呼ばれる突起物が存在しており,これが発生に伴ってさまざまな外の器官に変化していきます.
体節形成,器官の分化などについては,ショウジョウバエを使っての研究が進んでいて,体節を形成する分節遺伝子,各体節がどのような器官を形成するかを決めるホックス遺伝子などの調節遺伝子群が知られています.おそらくトンボにも同様の遺伝子群があって,体節形成と各体節に生じる器官の関係を決めているのでしょう.
体節がつくられていくと腹面中央に神経溝が出現します.昆虫の中枢神経は腹部の体軸方向に形成されるのです.脊髄ではなく腹髄です.この時期背中はまだ卵黄を背負っており (図6-1),組織や器官は十分に形成されていません.
*1.体節はヒトでも形成される.脊椎である.ヒトの脊椎は,頸椎7,胸椎12,腰椎,そして仙骨・尾骨から構成されている.例えば肋骨は,胸椎1番目から胸椎12番目から出ると決まっている.また手足の分化に関わる遺伝子は,ホックス遺伝子群である.このように体節が形成され,その体節が肋骨や手足の出る位置を与えている.
図5.トンボ胚の体節模式図と実際の対応.(a) Anax papuensis の孵化前の胚 (Tillyard, 1917 より).(b) ギンヤンマの下唇前基節で,一対の付属肢は孵化時に癒合して下唇を形成する.(c) 陥入したムカシトンボの胚を取りだしたもの.Aodo (1962) を参照して描画.(d) 形成された体節の模式図.(e) ギンヤンマの成虫.特定の体節から出ている付属肢は特定の器官に変化していくが,その運命は体節が形成されたときに決定されている.顎節と頭葉は一体化して頭部を形成する.そして胸節は胸部,腹節は腹部を形成し,それぞれ昆虫の「頭・胸・腹」を形づくる.図版は.
さて,体節が形成された胚は,次に大きな胚運動を起こします.胚反転です.これは胚の頭部と尾部が逆転する運動で,植物内に置かれる細長い卵では,頭部が植物の表面の方に向いて,脱出しやすくなるための位置変換といえます.
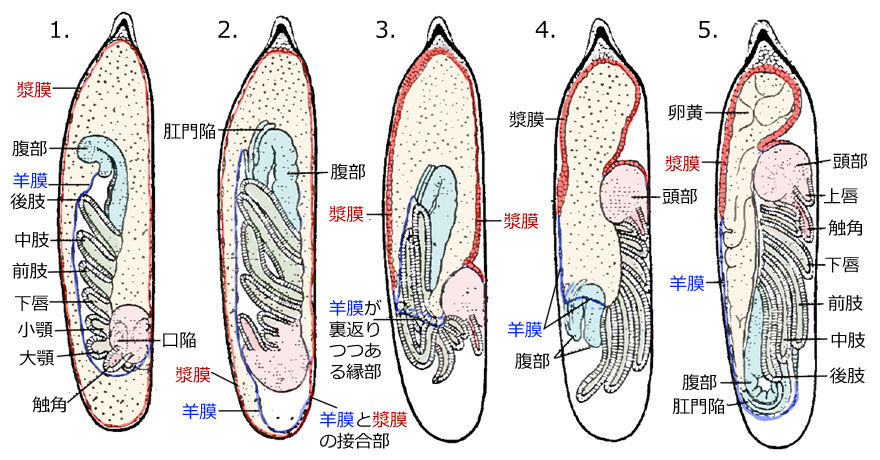
図6.Calopteryx virgo の胚反転.Brandt (1869) (in Tillyard (1917)) を改変.各器官名は,本来は「原基」語を後につけるべきだが,ここでは省略した.「口陥」は将来の口器,「肛門陥」は将来の肛門になる部位.ただしこの図ではこれらの形状ははっきりしていない.漿膜を赤色で,羊膜を青色で示した.この図は断面図であって,例えば漿膜も羊膜もこの絵の手前側や向こう側にも広がっている.もし断面のみを描けば「3,4の羊膜が裏返りつつある縁部」は出てこない.立体的に頭に描いて見ることが重要である.
胚の動きについて,図6の番号順に解説します.
- これは陥入が終わった胚で,体節や付属肢がすでに完成しています.陥入しながら羊膜 (青色で表示) が腹部側につくられていきます.この時点では羊膜は胚の側面から腹部を包んでおり,内部に羊水と呼ばれる体液を含んでいます.羊膜と漿膜 (赤色で表示) は頭部のあたりで接していますが,他は卵黄によって完全に分離しています.この時点でのイメージとしては,透明な袋 (漿膜) の中にもう一つ透明な袋 (腹側の羊膜と背中側は胚自身が袋の一部になる) があって,一部で接しているような状態を頭に描いてもらうとよいでしょう (欄外図7左).
- 胚の成長によって羊水の圧力が高まり,頭部が膨らんで隙間ができてきます.一方で漿膜は厚くなり収縮して,卵の後極付近に残っていた卵黄を卵の前極の方に引き上げはじめます.
- これら羊水の圧力の増加と漿膜に引き上げにより,羊膜と漿膜の接合部で両膜が破裂します.その破裂部位から頭部が外へ出て,卵膜に沿ってUターンするように前極の方に移動を始めます.先のイメージでいうと,接合部が開いて,羊膜+胚と漿膜でできた二重の靴下状になったという状態です (欄外図7右).この間にも漿膜は収縮し羊膜を裏返しながら前極の方に引き上げていきます.同時にその隙間にある卵黄も前極の方に移動します.その結果,胚部分は裏返る靴下の動きに合わせて反転するように出ていくことになります.ただ,この運動の力学に関しては漿膜の収縮力として説明していますが,胚自身の運動能力の可能性もあり,はっきりとしないということです (安藤,1996).
- 漿膜はさらに収縮して,羊膜との接合部で羊膜を引っぱり上げていきます.同時に胚はほとんど羊膜 (靴下) を裏返し終わって脱出しています.脱出し終わると,胚を包んでいた羊膜 (靴下) は完全に裏返ることになり,漿膜と一体となって卵黄を包む大きな袋をつくることになります.
- 胚反転が終わった状態です.胚は背中に裏返った羊膜と漿膜でできた袋に詰められた卵黄を背負うことになります.頭部は前極の方を向き,これは植物組織内に置かれた卵の出口の方です.こうやってやがては前極から孵化してくることになります.
この時点では胚の腹部側はかなり組織や器官がかなり分化していますが,背中側はまだほとんど何も形成されていません.それらが形成されるのは,このあと,
背閉鎖が生じてからです.背閉鎖とは,図6-5の胚の両側面から細胞が背中の方に広がっていき,背面の正中線でつながることです.これで卵黄は完全に胚の内部に取り込まれたことになります.
卵の形については次節でお話ししますが,トンボ科やエゾトンボ科などの球形に近い卵では,胚帯が一部しか卵黄内に陥入せず,前半身が表面に位置する
部分陥入型という状態で発生するものがあります.これらでは時に胚反転が生じないことがあり,その場合後極の方から孵化してくる場合が観察されています (新村,2001).
図6の中の
口陥と
肛門陥は盲管状の凹部を形成しており,将来口や肛門部になります.そしてそれらをつなぐように新たな細胞群が現れ,
前腸,
後腸がつくられます.さらにそれぞれの先に新たな細胞が現れ
中腸が形成され始めますが (中腸原基),これはつながることはありません.トンボでは中腸の中央部は卵黄細胞が分化して形成されます.中腸の完成は孵化後しばらくしてからになることがあり,この場合前幼虫や2齢幼虫の腹部に卵黄が見られることがありますが,これは形成されつつある中腸壁内部に含まれる卵黄です.(以上,安藤 (1996)).
以降,運動器官,神経系,感覚器官,気管系,消化器官,排泄器官,循環器官など,さまざまの器官が形成・完成され,あとは孵化を待つだけとなります.以上,大雑把に発生の流れを解説してきました.これらには,トンボの種によって若干の変化があります.また上では全く触れませんでしたが,胚が卵の長軸のまわりに回転するローテーションという動きが見られることがあります.ただしこれの役割ははっきりしていません.その他,詳しくは発生学の専門書にまかせたいと思います.
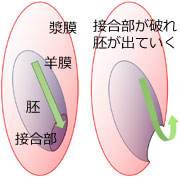
図7.羊膜と漿膜の動きのトポロジー.
左図では羊膜だけを閉じた袋のように描いているが,この内袋は羊膜と胚が一体となって形成されていることに注意する.また破れる部位は羊膜と漿膜の結合部なので,破れた後は二重の靴下状態となり,羊膜 (内袋) と漿膜 (外袋) の隙間にある卵黄は外に出ない.
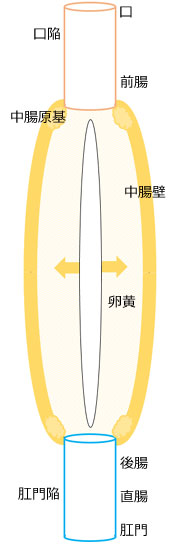
図8.口陥および肛門陥からそれぞれ生じる前腸と口および後腸と直腸と肛門,そして中腸原基および卵黄から形成される中腸,などのつながりを示す概念図.
トンボの生態学 4.卵と胚発生
卵の生態学的側面
前節では卵と胚の発生学的側面についてお話をしました.ここでは,卵の生態学的側面についてお話を進めたいと思います.なお卵の休眠の問題に関しては,
別の章で議論を展開していますので,そちらを参照してください.
Ⅰ.卵の形態と産卵様式
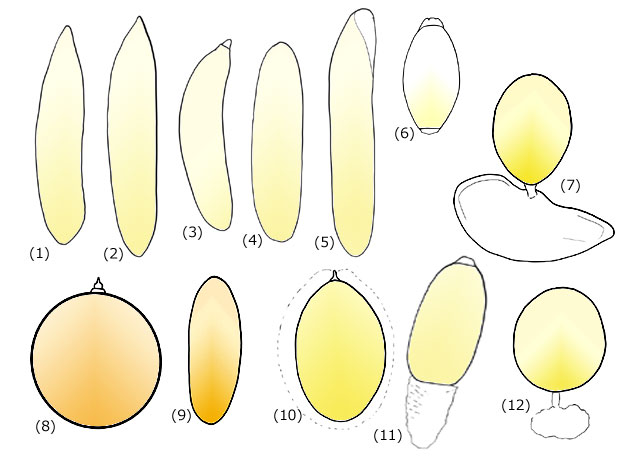
卵にはさまざまな形態のものがあります (図9).これらは産卵様式とある程度関連があります.ここでいう産卵様式というのは,まず植物内産卵か否か,植物外産卵の場合は産卵基質が何であるか,さらにその基質の上に置かれた卵の予想される運命などを指します.
図9.さまざまな卵の形状.(1) アオイトトンボ,(2) ハグロトンボ,(3) ホソミイトトンボ,(4) ムカシトンボ,(5) クロスジギンヤンマ,(6) オジロサナエ,(7) メガネサナエ,(8) リスアカネ,(9) コシアキトンボ,(10) タカネトンボ,(11) ウチワヤンマ,(12) オナガサナエ.拡大率は一定ではない.上を前極に描いている.卵の外部についているものはジェリー層である.
前節では細長いバナナ状の卵についてお話を進めてきました.このバナナ状の卵は,ほぼすべての植物内産卵の種に共通した形状です (図9(1)~(5)).後極から植物組織の中に押し込まれ,前極が植物表面の方を向いています (図10).ですから胚反転によって頭部を前極の方に向ける必要があるということになります.均翅亜目の全種とヤンマ科の卵がこれに該当します.ただムカシヤンマの卵は,産卵管で苔などの間に産み込まれますが,バナナ状とまではいかず長楕円体をしている点で少し異なっています.
図10.植物組織内に産み込まれた卵 (コバネアオイトトンボ).左は産卵口で植物の表皮によってふたができている.右はその部分をカミソリで切り開いた状態.
植物組織外の基質に産卵する卵は概形として楕円体をしています.これらの卵には,さまざまの付属物がついています.特に外卵殻が変化したジェリー層は,通常水分を吸収して粘着性を持つようになり,卵を基質に付着させる役割を果たしています.例えば植物上産卵をするコフキトンボ,アオビタイトンボ,コシアキトンボの卵には粘着性のあるジェリー層があり,卵が植物表面に貼りつきます (図11).
このうちコフキトンボとアオビタイトンボはふつうわずかに水面下に位置する基質上に卵を置きますが,コシアキトンボはわずかに基質の水面上の位置に卵を貼り付けています.前二者は卵と水がふれあうことが保証され,ジェリー層が水に触れて粘着性を持つことができますが,コシアキトンボの卵はこのままでは水に触れることができません.しかし何度も観察を続けていると,コシアキトンボは水に濡れている産卵基質を選んで産卵していることが分かります (欄外図12).このように詳細に観察すると,トンボの行動には非常に細かい戦略があることが見えてきて興味がそそられます.なお植物上産卵をするある種のトンボのジェリー層は,水に触れなくても粘着性を持つものがあることが知られています (Corbet, 1999).
図11.植物表面上に産み付けられた卵.左はアオビタイトンボで右はコシアキトンボ.コシアキトンボの方は産み付けられた直後の白い卵がしばらく時間が経った褐色の卵に重なって産み付けられている.見にくいが,卵のまわりにジェリー層があるのが見える.
トラフトンボはジェリー層が大きな役割をしている卵の一つです.トラフトンボはカエルのような「ひも状」の卵を産むことでよく知られています (図13(5)).トラフトンボは,交尾が行われた後,交尾態のまま放卵するポイントを探しに池を訪れます
(1).放卵のポイントが決まるとオスはメスを放します.そしてメスはすぐに打水します
(2).その後メスは周辺樹林や草地に飛び,産卵弁と第9,10腹節の間に卵塊を形成しはじめます
(3).卵がばらばらにならずに塊をつくるのは,直前の打水によって水を産卵弁あるいは生殖口に含ませ,それによって卵のジェリー層が「少し」膨らみ粘着性を持つようになるからと推測されます.卵塊が形成されると,メスは先ほどの放卵のポイントを訪れ,水面をこするように移動して放卵します
(4).これはできた卵塊を引き延ばしてひも状にするための行動だと思われます.しばらくすると放卵ポイントには,ひも状の卵が沈積物に絡まっているのが見られます
(5).

図12.コシアキトンボが卵を貼り付ける位置が,水に濡れている場所であることがよく分かる写真.水から突き出した石に放卵するコシアキトンボは,石が水に濡れている場所を選んで卵を貼り付けている.他の産卵基質の場合も,卵を貼りつけている基質は水に濡れている.
図13.トラフトンボの卵塊と産卵様式.(1) 交尾を終えたペアは交尾態のまま放卵するポイントを探しに池を訪れる.(2) 放卵するポイントが決まるとオスはメスを放しメスは一,二度打水する.(3) その後メスは池周辺に止まって卵塊を形成する.(4) 卵塊を形成し終わったメスは先の放卵ポイントを訪れ,水面をなでるようにして卵を放す (1/6秒間隔,4コマの合成写真).(5) 産み落とされた卵はジェリー層が膨らみ互いに接着してひも状の卵塊となる.
トラフトンボのひも状の卵は,「ひも状のジェーリー物質」の中に卵が入っているのではなく,個々の卵のまわりの膨らんだジェリー層によって,それぞれの卵がくっついてできているものです (欄外図14).このジェリー層に粘着性を持たせるために,卵塊形成前に一度打水する必要があると考えられます (図13(2)).そして水面をなでるように産卵するのは,卵塊を引き伸ばすためであり,引き伸ばされた卵塊のジェリー層はさらに水を吸って膨らみ,最終的にひも状の卵になるのでしょう.
卵についている付属物は生態学的に興味深いことが多いものです.次はウチワヤンマを見てみましょう (図9(11)).ウチワヤンマの卵には,後極に糸を巻いたような付属物があります.これは水に触れるとほどけ何本もの細い糸になって広がります.ウチワヤンマは6月ごろに植生があまりない池によく現れ,水面に浮かんだ浮遊物に卵を絡みつけます.浮遊物といっても植生のない池ではあまり浮かんでいる物体は多くはありません.メスは数少ないそういった浮遊物に腹端を打ちつけて卵を貼り付けます (図15).次々とメスが同じ浮遊物に産卵するので,浮遊物はこの卵糸がからみついてすごい状態になっています (図16).
図15.浮遊物に貼り付けるように産卵するウチワヤンマのメス.後極の粘着性のある卵糸がほどけ,浮遊物に絡みつき,腹端に残された卵と糸でつながっている.拡大してみると卵が糸に付着しているのが分かる.
図16.浮遊物の拡大写真.それ以前に産み付けられた糸がたくさん絡みついていて,そこかしこに卵が見える (矢印).
図9に示された卵のジェリー層の中でも,メガネサナエの卵後極についている吸盤状のそれ (図9(7)) は,見ただけでその役割が分かりそうな付属物です.
通常メガネサナエは,大湖の波打ち際や,そこに流れ込む河川で産卵します.はじめはこの吸盤状の付属物は反転していて卵のまわりに広がっていますが,水に浸かると外に広がって吸盤状になります.そしておそらく底の石や砂などに吸着するのだと考えられます.
飼育条件下ではそのようすが撮影されています.そうなることで,波打ち際では波によって浮き上がることや,川では流水に流されるのを防いでいるのでしょう.

図14.形成中のトラフトンボの卵塊.よく観察すると,最初から細長いジェリー層が卵を包んでいるわけではないことが分かる.個々の卵のまわりにうすくジェリー層がついているだけである.
図17.大湖の波打ち際で放卵するメガネサナエ.
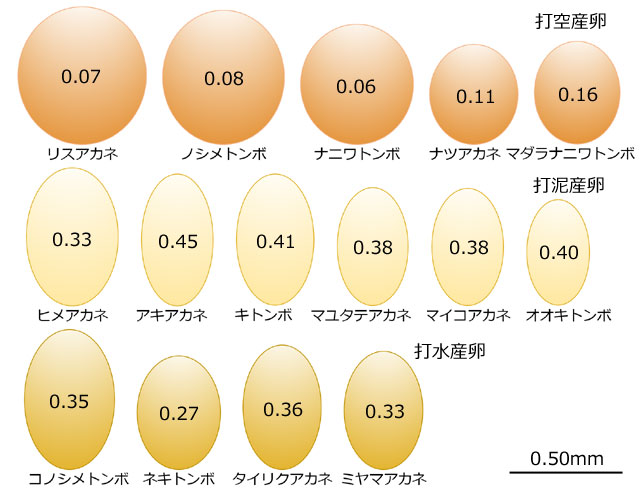
では次に,産卵様式と卵の形について見てみましょう.これに関しては,兵庫県産のアカネ属を使った観察と考察を新村 (2001) が行っています.アカネ属のトンボは,打空産卵,打泥産卵,打水産卵と,いろいろな様式の産卵を行いますので,比較検討しやすい近縁の一群です.この研究にはアカネ属15種の卵の長径と短径が記されていますので,卵の形と大きさを楕円体として模式的に表示してみました (図19).
図19.兵庫県産アカネ属の卵の大きさ比較と,長軸に沿った断面楕円の扁平率 (=1-短径/長径)を表示した.扁平率は真円に近いほど0に近づく.打空産卵するトンボの方がより真円に近いことが分かる.データは新村 (2001)より.
これを見てまず気づくことは,打空産卵をする卵は球形に近い形状をしているこです.これら5種は,いずれも水際の,水のない陸上に卵を落とすような産卵様式です (図20).またこれらの卵を取りまくジェリー層は,いずれも産卵後しばらくは少ないことが観察されています (新村,2001).
図18.メガネサナエ卵のジェリー層による石などへの吸着.
図20.ナニワトンボの産卵場所.水落をして底が露出した池の縁で,水際から離れた陸上に卵を落とす.
これらのことから推定できることは卵が転がりやすいということで,図20のような場所に卵を落とした場合,落ち葉や石の上を転がってその隙間に入り込み,うまく隠れてしまうことでしょう.またジェリー層はやがて出てくるということですが,この場合は,上の植物上産卵の卵と違って,まわりの土などを身にまとって姿を隠すことに生態的意味があると思われます.転がって水の中に到達するというのは,陸上に産卵する意味からは考えにくいでしょう.
打水産卵,打泥産卵をするアカネ属の卵はどれも大差なく,扁平率が0.30~0.40の,楕円体です.これらは基質の中に置かれるので転がるということとはまず無関係です.ジェリー層も多少は存在するようですが,特徴的なことはありません.唯一ミヤマアカネのジェリー層に関して,「産卵後数時間で卵を取り巻き,他の物質に強力に付着する」と書かれています (新村,2001).ミヤマアカネは流水に産卵しますので (図21),卵が流されないように,ジェリー層がこのような性質を持っているのかもしれません.
図21.ミヤマアカネの産卵場所.流水で打水産卵する.卵が大きく流されないためには,ジェリー層の固着力は役に立つと思われる.
最後にもう一つ,ここでは推論しかできませんが,幼虫の流下説について考えてみましょう.例えばヒメサナエは河川性のサナエトンボで,羽化場所が繁殖場所よりかなり下流よりになっていることで知られています.一般的には,この現象は幼虫が積極的に,あるいは受動的に,川を下ることによって実現されていると考えられています (例えば新井,2001).ヒメサナエは,欄外図22のように流れの速い場所の真ん中に打水して放卵する性質があります.このとき,もし粘着性のジェリー層が (少なくともある程度の期間) 出現しなければ,卵の状態で流されている可能性があります.新井 (2001) では,産卵水域で幼虫が採れないとも書かれています.つまり少なくとも流下移動のいくらかは,卵の流下によって生じているといえるかもしれません.
以上のように,卵の形およびジェリー層の性質とそれが現れるタイミングが,生態学的に重要な意味を持つことが分かります.卵そのものは動けませんが,それだけに適切な位置に自分自身を置くための仕掛けが備わっているように思います.
Ⅱ.卵の捕食者
トンボ卵は,魚や貝などによる捕食,寄生峰による胚の捕食などを受けます.打水産卵は卵を直接水中にばらまくため,その卵は特に魚類の捕食を受けやすいと思われます.実際ミヤマアカネで,打水する度に打水したあたりの卵に群がる仔魚を観察したことがあります (ビデオ1).
ビデオ1.ミヤマアカネが打水産卵したあと,その卵に群がる仔魚たち (6秒,音声なし).
ミヤマアカネは,図21のように,通常もっと流速の速いところや石の間の小さな水面に放卵する.これはたまたまの産卵失敗と思われる.
これはある意味当然のことで,卵が餌として小さすぎる大きい魚は別として,小さな魚にとっては栄養価の高い卵はよい餌になることは間違いありません.しかしトンボの方も食べられないような戦術を進化させているはずです.もっとも基本的な戦術は,小さな魚がいないところに産卵するというものでしょう.一番ふつうに見られるのは,水中の魚が確認できるような,水が濁っていない場所で産卵することです.観察経験から考えても,全く水中の様子が見えないような濁った場所では,多くのトンボは打水産卵することが少ないように思えます.それ以外にもさまざまな工夫が見られます.
例えば, ネキトンボ,コノシメトンボ,キトンボなどは仔魚のいないような深い場所を選んで放卵していることがあります.ヒメサナエ (図21),アオサナエ,オナガサナエ,ヤマサナエ,ミヤマサナエなどは,川の流速の速い場所を選んで放卵しています.ショウジョウトンボなどは水草のすぐ上の場所を選んで卵が見つかりにくくなるようにしています.オオシオカラトンボ,シオヤトンボ,ヨツボシトンボなどは,打水して水を飛ばし,打水場所から離れたところにある物体に卵を付着させる,あるいは落とすような産卵をしています (図23).

図22.ヒメサナエの放卵の瞬間.ヒメサナエは流れの速いところに産卵する.
図23.打水産卵,水面上での空中産卵を行う種の捕食を避ける産卵戦術.(a) タイリクアカネ:水中の見通しが良い水が透明な場所での打水産卵.(b) ネキトンボ:深い池の中ほどでの打水産卵.(c) オナガサナエ:流速の速い場所での空中産卵.(d) ショウジョウトンボ:水草が繁茂する場所への打水産卵.(e) オオシオカラトンボ:卵を含む水滴を水際の植物に向かって飛ばす打水産卵 (飛水産卵).(f) ヨツボシトンボ:水滴を打水地点から前方へ飛ばすような産卵.これらいずれも,仔魚にねらわれにくいような場所,あるいは攪乱するような方法である.
さらに筆者は興味深い観察をしています.コフキトンボとコシアキトンボは,いずれも植物上産卵を行うトンボです.コフキトンボは,観察した限りは,わずかに水面より下に沈んでいる基質に卵を貼り付けており,産卵後の卵は水面よりわずかに下に置かれています.対してコシアキトンボは,こちらも観察した限りでは,水面よりわずかに上に出ている基質に卵を貼り付けています (図11右).
ある日,コフキトンボを観察しているとき,過ってコフキトンボの産卵した水草の葉を蹴ってしまい,葉が卵とともに水面に顔を出しました.そのときは特に気にしませんでしたが,しばらく経ったあと,その場所に戻ってみると,モノアラガイやアメンボが水面に出た卵をおいしそうに食べていたのです.水面下に置かれた卵がこのような捕食を受けるのを観察したことはありません.またコシアキトンボの卵がモノアラガイやアメンボに食べられているのを見たこともありません (図24).
図24.コフキトンボの産卵と卵が置かれる位置.(a) コフキトンボは,水面よりわずかに下にある基質に卵を貼り付ける.(b) わずかに水面より下に位置する葉の上に接着しているコフキトンボの卵.(c) その卵を水面上に出したらモノアラガイやアメンボが集まってきて,全部食べてしまった.
この場合捕食者は魚ではありませんが,この対比には非常に不思議な何かを感じました.水中に置かれているコフキトンボの卵には何か捕食を妨げる化学物質のようなものが出ているのでしょうか.また水面上に置かれているコシアキトンボの卵にも同様の何かを感じます.またヒメクロサナエが無造作に湿った基質の上に卵を置いていることがありますが,これも捕食されないのか不思議です (欄外図25).こういったしくみについて,手段を持っている方々によって,もっと調べられたら面白いと思います.
トンボの産卵方法を,単に,打水,打泥,空中,打空,等々のトンボの動作や産卵基質との関係で分類するだけでなく,卵を捕食する生物との関係で見ていくと,興味深いトンボの産卵戦略の詳細が見えてくるでしょう.なお複数の産卵様式を持つトンボは,自身の有する複数の戦術を産卵場所の水環境によって使い分ける戦略を有しているということで,なかなかの優れものといえるでしょう.

図25.湿った落ち葉の上に無造作に置かれたヒメクロサナエの卵.
卵の捕食者として,あまりにも小さくて気づかれないですが,それなりに大きな影響を及ぼしている昆虫が,卵寄生蜂です (欄外図26).主に植物内産卵や植物上産卵される卵に寄生することが多いようです.新村 (2004) に,卵寄生蜂の観察結果が報告されています.そこにはタマゴヤドリコバチ科,ホソバネヤドリコバチ科,ヒメコバチ科,タマゴクロバチ科の卵寄生蜂の観察結果が記されています.宿主としては,アオイトトンボ科,モノサシトンボ科,イトトンボ科,ムカシトンボ科,ヤンマ科などが挙げられています.
トンボの卵内に産みつけられた寄生蜂の卵は,やがて孵化して幼虫となります.幼虫はトンボの卵黄や胚を食べて成長します.そしてそのまま卵内で蛹化することがふつうのようですが,幼虫が卵から脱出しして,卵の外で蛹化するものもあるそうです.前者は卵内で羽化して,大顎で卵殻を砕いて出てきます.出てきた寄生蜂は,オスの場合そのあたりをうろついて,他の卵から出てきたメスを見つけて交尾するそうです.なお,通常1卵に1頭の寄生蜂が入っています.複数入っている場合は,卵内に顎の残骸が残っているのが観察されることから,寄生蜂どうして争いが起き,最終的に1頭になると考えられています (以上,新村,2004).しかし Corbet (1999) には,「大きな卵ほど多くの個体が生まれる」との記述があって,このあたりはさらに調査を進める必要があるでしょう.
さらに海外での観察から,いくつかの興味深い点が指摘されているので,二つほど紹介しておきましょう.
カナダのケベック州において,アオイトトンボのガマの葉に産卵されていた卵について,葉の先端に近い方では寄生率が95%だったのに対し,茎に近い基部の方では45%であった (Laplante, 1975) という報告があります.95%というのは驚くべき高さです.個体数が激減した希少種にこのようなことが起きると,一気に個体群消滅への道を歩んでしまうでしょう.
また,植物内産卵をするトンボが,水面下に腹部を沈めて産卵するのは,卵寄生蜂の寄生を防ぐためである (Corbet, 1999) という考察があります.普段何気なく見ているアオハダトンボなどの産卵にも,こういった戦略があるのかと思うと,その産卵姿勢に,より深い理解が進むような気がします.また潜水産卵を行うトンボは,オスに邪魔されないためというのもあるかもしれませんが,卵寄生蜂を避ける手段という見方も成り立ちます.

図26.卵寄生蜂,ホソバネヤドリコバチ科
Polynema 属.Miall (1895) in Tillyard (1917).
カワトンボ科の卵に産み付けられたもので,卵内で孵化すると数日の内に卵の内容物をすべて食べてしまい蛹化,その後10-12日で羽化した.翅を使って
泳ぎ,スイレンの葉に産み付けられた別のカワトンボ科の卵を探した.
図27.腹部を水面下に沈めて産卵するトンボたち.左:セスジイトトンボ:腹部を水面下に沈め,さらに葉の裏側に産卵している.卵寄生蜂に見つかりにくくするための戦術と見ることができる.右:ミヤマカワトンボ:こんな浅いところでもわざわざ潜水して産卵するのは,オスの干渉を避けるためもあるだろうが,ひょっとしたら,卵寄生蜂を避けるためかもしれない.
筆者は卵寄生蜂に関しては門外漢なので,実物写真を見たい方は,
美しい写真とともに解説されている「近畿のトンボ雑記」のページをご覧ください.卵寄生蜂が個体群動態にどの程度の影響を与えているか,個体数が激減したトンボを絶滅させるほどの圧力になるのか,まだまだトンボについて知られていないことが多いです.
Ⅲ.卵の生存率
最後に,卵の生存率について少しふれておきましょう.Corbet (1999) は,卵の生存率を次のように定義しています.
(卵の生存率)=(孵化した2齢幼虫の数)/(産下された卵の数)
ここで2齢幼虫としているのは,植物内産卵の細長い卵では,前幼虫が完全に卵から脱出する前に,2齢幼虫が脱皮して出てしまうことが多いからです.詳細は後で議論することにしましょう.
このように定義したときの生存率に影響を与える要因は,一つは温度・乾燥・酸素不足などの物理的要因による死亡,もう一つは捕食・寄生・病気などによる死亡,そしてもう一つが受精率です.受精率というのは生存率という概念には似合わない気がしますが,受精したかどうかは外観からはすぐには分かりませんし,病気,酸素不足,卵の不具合などによる初期の発生進行停止とも区別がつきませんので,この定義では考慮しておく必要があります.
日本のトンボでは,曽根原 (1982) がオオトラフトンボの孵化率を報告しています.オオトラフトンボはトラフトンボと同じようにひも状の卵を産みます.それによると,孵化率は68.8%-96.7%となっています.孵化しなかった卵は,そもそも胚発生が見られなかった卵で,未受精卵,または卵紐内での酸素不足による死亡の結果であった可能性があると述べられています.
トンボの生態学 4.卵と胚発生
卵期間と温度
ここでは卵期間に影響を与える温度要因について見ていくことにしましょう.トンボの卵期間は種によって異なります.特に休眠性の卵を産む種は一般に卵期間が長くなります.卵を飼育していると,採卵後,孵化までの時間にばらつきが生じ,卵期間がある範囲の広がりを持つことがふつうです.
Bennett & Mill (1995) は
Pyrrosoma nymphula という非休眠性の卵を産むイトトンボを使って,卵の産下から孵化までの日数を調べています.245卵が得られ,15~24℃の範囲で飼育され,孵化率は75.1%でした.孵化は22日目から始まり56日目に終わり,その中央値は26日後でした.その際特徴的だったことは,孵化日の上限に近づくほど孵化個体数は漸近的になるということでした (欄外図28).Corbet (1999) は,このように上限が漸近的になることは一般的なことであって,種ごとの卵期間を比較するにあたっては,上限値はあまり意味を持たず,最短卵期間が比較の意味を持つと述べています.
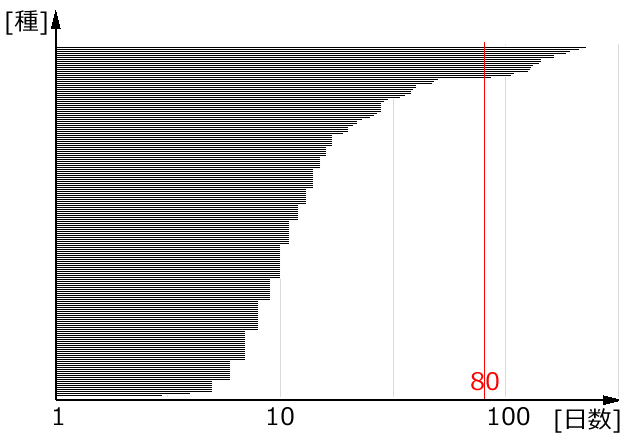
そこで,吉田 (2014) がまとめた卵期間のデータをもとに,そこに記載のある175種の最短卵期間 (の常用対数値) を長い順に並べて表示したのが図29です.
図29.日本産トンボ175種の飼育による最短卵期間を長いものから並べてくラフにしたもの.吉田 (2014)を参照した.
グラフによると,80日を越えないものと越えるものとの間に少し開きが見られる (36日間).一般的に80日以上の卵期間を持つものが内因性休眠卵と考えられている.実際,図で80日以上の最短卵期間を持つ種は,ルリボシヤンマ属,アオイトトンボ属,アカネ属など,すべて
秋季種である.最短卵期間が80日未満でも,経験的な判断で卵越冬 (つまり休眠卵を産む) していると考えられる種も,多く含まれていることには注意したい.例えばアキアカネは,飼育卵期間の範囲が39~202日間となっており,非常に範囲が大きい.飼育下でこのように短期間で孵化に至る卵の存在に関しては,アキアカネの卵休眠の「浅さ」が関係しているという考え方がある (上田,1993).
図29の中には休眠性の卵を産むものも含まれています.特に内因性の休眠卵は,温度と卵 (胚) の成長に関して相関性がなく,いくら適温化に置かれても発生が全く進まないという特徴があります.ここでは卵期間 (胚発生の完了までの期間) に与える温度の影響を見るので,非休眠の卵 (
直接発生卵ともいう) についてのみ考えていくことにします.
直接発生卵は,適切な飼育環境下に置かれた場合,変温動物一般の成長についてよく知られている,
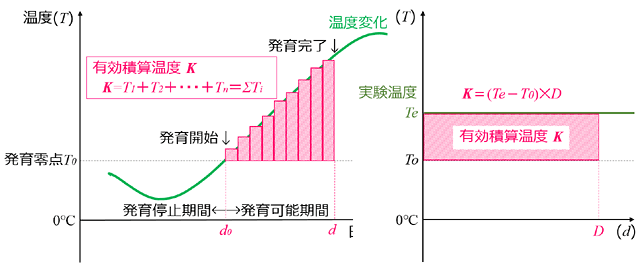
有効積算温度の法則にしたがって胚発生が完了し,孵化するようです.簡単に説明すると,まず,直接発生卵には,ある温度以下であると発生が進まなくなる限界の温度があって,それを
発育ゼロ点といいます.それ以上の温度になると発生を始めることになるので,発育ゼロ点以上の温度が発育にとっての有効温度となります.そして発育が完了するのは,発育開始から毎日の有効温度を積算していったものがある一定値に達したときである,という一般論が成立することが知られています.これを有効積算温度の法則といいます.
図30.有効積算温度の概念図.仮想的なトンボ種の卵について,左図には各日の有効温度の代表値 (通常は平均温度) を棒グラフで表示した.各棒グラフの面積は,1日X温度 となり,単位は[日・度]となる.発育開始から発育完了までの棒グラフの面積を合計したものが,有効積算温度となる.実験的には,右図のように恒温下で飼育が行われることが多い.この場合,発育ゼロ点も不明であるので,有効積算温度と発育ゼロ点の両方を実験から求める必要がある.
実際にこれらを厳密に測定した結果が,アキアカネについて得られています.アキアカネは休眠卵を産みますので,ここでの話にあわないように思われますが,越冬後であれば休眠が消去されており,いわゆる温度依存の成長のステージに入っています.ただし,通常アキアカネは越冬前に胚反転を終えており,ここでの測定は,胚発生の一部分について行われていることは承知しておく必要があります.
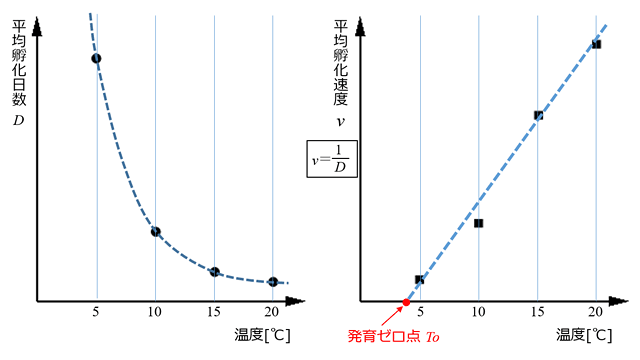
神宮字ら (2006) は秋田県八郎潟で10月18日に採集されたアキアカネの卵を使い実験を行いました.野外で越冬させたあと,4月21日に実験室に持ち込み,日長条件 LD14:10 で,20℃,15℃,10℃,5℃,および暗黒下で20℃という恒温下において飼育し,ばらばらに孵化してくる卵の平均孵化日数を求めました (図30右の
D に相当).平均孵化日数と飼育温度の関係は,一般的には図31左のように双曲線的な曲線を描きます.そこで実験的処理としては,
D の逆数をとり,これを孵化速度と定義してグラフを描くと,一般的には直線になります (図31右).
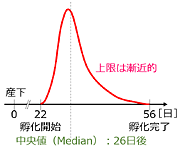
図28.
Pyrrosoma nymphula の孵化曲線.(Bennett & Mill, 1995) の記述に基づいて筆者が描画.
図31.実験的に求めた発育ゼロ点と友好積算温度 (神宮字ら,2006のデータを参照).日長条件 LD14:10 の4つの恒温条件下の結果をプロットしたもの.左は平均孵化日数と温度の関係で,双曲線的な曲線を描いている.右は平均孵化速度と温度の関係で,平均孵化日数の逆数 (1/D) と温度の関係.これは経験的に直線になる (Pilon & Masseau, 1984 参照).右の回帰直線の方程式を,v=at+b とおくと,v=0 の時 (つまり発生が止まっている) の温度 To が発育ゼロ点となる.また 傾き a の逆数が有効積算温度になる ※2.なおこの方法は,桐谷 (1997) の論文をもとにしている.
神宮字ら (2006) は,図31のようにして発育ゼロ点および有効積算温度を求め,それぞれ 4.9℃,および 54.6日度 を得ています.
※2.傾き a の逆数が有効積算温度になる理由:
v=at+b --- (1)
発育ゼロ点を To とすると,発育が進まないということは発育速度 v が v=0 ということなので,
t=To のとき v=0,
これを代入すると,
0=aTo+b
aTo=-b
∴To=-b/a --- (2)
次に(1)の両辺をavで割ると
1/a=t/v+b/av
ここに,v=1/D を代入
1/a=tD+bD/a
1/a=(t+b/a)D
ここに(2)を代入すると
∴1/a=(t-To)D
tを実験温度Teに置き換えると
∴1/a=(Te-To)D
となって,右辺は図30右の有効積算温度 K に一致する.つまり,a の逆数は有効積算温度となる.
ところで,Pilon & Masseau (1984) の予備的考察によると,直接発生卵は上述のように発育速度が温度と直線的な関係が成り立ちますが,これは発育最適温度と呼ばれる範囲内でのことだと述べています (欄外図32).まず発育が可能な温度上限,下限が存在します.それぞれ発育有効温度上限 (Tmax),発育有効温度下限 (Tmin) と呼ばれています.発育最適温度はこの範囲内にありますが,それぞれの温度より若干低い温度および高い温度の範囲にあるのが一般的です.発育最適温度の範囲を超えると,発育速度は図31右のような温度との直線関係から外れ,さらに発育有効上限温度・下限温度を超えると発生が完了しなくなり,死亡率もかなり高くなります.また発育が完全に停止する低温の閾値が存在します。彼らはこの温度を「閾値 Threshold」と表現していますが,これは発育ゼロ点と考えられます.
彼らの研究では,いくつかのイトトンボ科について,これらの温度が測定されており,例えば,ルリイトトンボ属の Enallagma ebrium では,To=5.0℃,Tmin=12.40℃,最適温度の範囲 22.5-30.0℃,Tmax=32.5℃,となっています.ただこれらの値はこの種における厳密な値ではなく実験で設定された温度 (恒温) である点は注意しておく必要があります.
桐谷 (1997) は,主として日本に分布する430種の,昆虫,ダニ,線虫の,総計約600の報告をもとに,発育ゼロ点,有効積算温度の概括的な分析を加えています.これによると Pilon & Masseau (1984) と類似のことが記されており (欄外図32),通常10-35℃の温度領域で任意に設定した (つまり最適発育温度内に設定) いくつかの温度区 (恒温) で昆虫を飼育して実験を行うように書かれています.また発育ゼロ点 To の推定が,直線部分の外挿による (図31右) という技術上の問題点も指摘されています.なお,トンボ目については,上田 (1993) によるアキアカネ卵の発育ゼロ点が10℃であることが記載されているだけです.
齊藤ら (2017) は,仙台市内の個体群を使って,神宮字ら (2006) に準じてアキアカネの発育ゼロ点などを求めていますが,5℃の恒温下では孵化が進まなかったと書かれています.おそらくこの個体群においては,5℃は,発育最適温度範囲,または発育有効温度下限を下まわっていたのでしょう.

図32.卵の発育可能な温度範囲.
Tmax:発育有効温度上限,
Tmin:発育有効温度下限,
To:閾値 (発育ゼロ点).
この発育と温度の関係が正しいとすると,発育速度と温度の関係が直線関係にならない部分 (
Tmin と発育最適温度の間の白の部分) が存在するので,図26で求めた発育ゼロ点は理論上の値ということになろう (桐谷 (1997) も参照).Pilon & Masseau (1984) に基づいて筆者が描画.
トンボの生態学 4.卵と胚発生
孵化
卵の時代の最後は孵化です.上で述べましたが,孵化は,卵殻が破れて前幼虫が出始めたところから始まり,2齢幼虫が完全に脱出し終わった時点で終了します.本サイトでは前幼虫を1齢と数えています.Corbet (1999) は手元の資料の6割程度は前幼虫を1齢としているということを述べています.さまざまな文献を読むときに,前幼虫と齢数の割り当てがどうなっているかを確かめることが肝要です.
さて,孵化の実際を見ていくことにしましょう.これはトンボの分類群や産卵される場所によって,若干の相違があります.ここでは Tillyard (1917) の観察をもとに Anax papuensis の孵化を見ていきましょう (図33)*3.彼はこの観察を非常にていねいに行っています.最近は顕微鏡を使ってじっくりと観察した記録というものがあまり発表されないので,この古典的な観察記録は重要だと考えています.そこで少し要約して詳しく紹介しましょう.
Anax papuensis の卵は,ヨシなどの茎の水面すぐ下の位置に産み付けられます.孵化前の胚はすでに胚反転を終えており,頭部が前極の方に向いています (図33A).大顎,小顎などの口器はすべてはっきりと確認できます.下唇は1対の付属肢の基部だけが融合した状態で,脚と同様に後方を向いています.腹部の後端は腹側に湾曲して前方を向いた状態です.背気管幹はまだ液体で満たされています.中腸は卵黄で形成されつつある状態のため (図8),他からはっきりと区別できます.
*3.Corbet (1999) は,ギンヤンマ族 tribe Anax には,他のヤンマ科に存在する「卵歯 (頭楯のすぐ上にある卵殻破砕器)」がないと記されているが,日本産のギンヤンマにはあるという.渡辺 (1997) のギンヤンマ卵の写真においても卵歯は明瞭に確認でき,記載もある.したがって,ここでの説明では「卵歯によって卵殻を切る」という表現がないことに注意したい.
孵化の2,3日前になると,心臓が拍動し始めます.最初は1分間に約30回程度の不規則な脈動が見られますが,やがて1分間に80回以上になります.頭部とペディセルの間には大きな小胞 (図33A, Bの head vesicle) が形成されます.孵化直前になると,頭部に特徴的で独特な拍動器官である頭心臓が口と大動脈の間に現れます.これはおそらく食道内の一時的な構造物であり,この拍動によって頭部が大きく膨らんで,ペディセルを内部から押し上げ,卵殻が破れて,前幼虫が流れるように出てきます (図33B, C)*4.
*4.この部分を安藤 (1996) は,「...頭心臓が出現し,前幼虫は卵内の液を嚥下する.その結果,体内圧が高まって卵の先端部が膨張し,前幼虫はその部分を卵歯で切り開いて欄外に脱出する.」と,ギンヤンマでの観察結果を記述している.
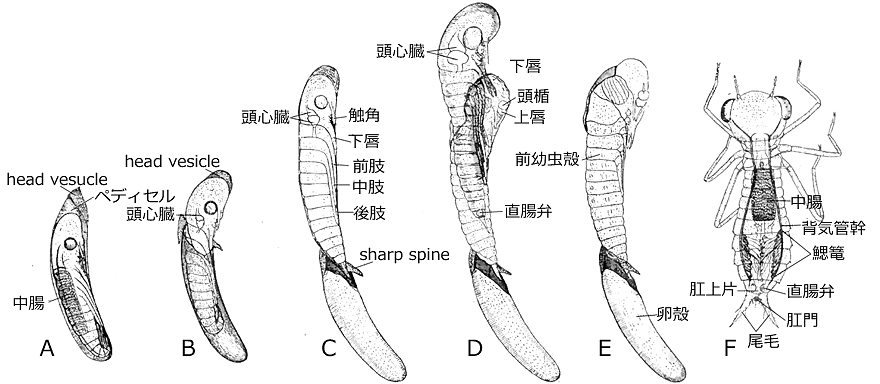
図33.Anax papuensis の孵化の順序.(Tillyard, 1917を改変).A:胚反転・背閉鎖を終えた孵化前の卵.B:卵殻が破られ孵化が始まった.C:sharp spine によって卵殻に引っかかり,それ以上脱出しなくなった前幼虫.D:その後直ちに2齢幼虫が出てくる.E:卵殻と前幼虫の脱皮殻.F:脱出した2齢幼虫.
前幼虫全体は前幼虫鞘と呼ばれる,薄いキチン質の鞘に覆われています.この鞘は脚などを外側からぴったりと押さえつけるように密着しており,触角,下唇,前肢,中肢,後肢などは,腹面に密着させられた状態になっています.これはあくまで私の主観ですが,この鞘は,いわゆる完全変態をする昆虫 (例えばカブトムシ) の蛹の皮を連想させます.この前幼虫鞘は尾部先端で鋭い棘 sharp spine の形になっており,これが卵殻に引っかかって,前幼虫は卵から完全に脱出せずにとどまります (図33C).
この種では前幼虫期はわずか3-20秒程度しか続かず次の脱皮が始まります.その過程で,頭心臓は著しく大きくなり,head vesicle 内の血液を頭部の背側の方に送り込みます.そしてこの背側の部位が大きく膨らむことで,前幼虫鞘が頭部から胸部にかけて背中側が破れ,2齢幼虫が出てきます (図33D).頭心臓はやがて見えなくなります.
3節からなる下唇は,先端の2節が次第に折れ曲がり,それぞれ下唇前基節と下唇側片となり,下唇が完成します.これでやっと採餌の体勢が整ったことになります.同時に気体が,中腸付近から背気管幹を通って広がっていくのが見えます.この気体は二酸化炭素であり,まもなく鰓篭が機能し始めると,空気と置き換えられていくようです.この2齢幼虫には単眼がなく触角も3節でできているようです.
以上,Tillyard (1917) の観察を紹介しました.多くのトンボの孵化過程は.この Anax papuensis と同様ですが,卵が置かれる位置によって,若干のバリエーションがあります.
例えば日本のトンボでは,アオヤンマの観察があります.フトイの,水面より高い位置の内部に産み落とされた卵 (欄外図34) から孵化した前幼虫は,卵殻から完全に脱出し,フトイの産卵傷口から外に出て,水面に落ち,そこで2齢幼虫に孵化します (井上ら,1981).水面に落ちなかった場合は跳ね回って水面にたどり着いてから,2齢幼虫に脱皮します.実験的に水面に到達しないようにした前幼虫について,14時間後に水面に浮かべた場合でも2齢幼虫に脱皮したと書かれています.通常水は低い位置にありますから,跳ね回っている間に水にたどり着く可能性は高いでしょう.
また Tillyard (1917) には,日本のオオアオイトトンボと同じように樹皮に産卵する Lestes viridis について,「前幼虫は鞘に包まれた状態で出現し,<中略>,体を強く曲げることで茎から飛び出し,通常は水中に落ちます.
しかし地面に落ちた場合は,川に向かって傾斜した地面を利用して勢いよく跳ね回り,最終的に水中に飛び込みます.水中では、背側を下にして浮かびます.2齢幼虫の出現は Anax 属の場合とほぼ同じです.」と,Pierre (1904) の観察を引用しています.
一般に,水から離れた位置で孵化する場合は,このように前幼虫の期間が長いのがふつうです.
無事孵化した2齢幼虫は,水生動物を構成する一員として,厳しい生存競争のまっただ中に放り込まれていきます.

図34.アオヤンマがフトイに産卵する.フトイの茎の中空部にはたくさんの卵が詰め込まれているという (井上ら,1981).
参考文献
青木典司,1997.神戸のトンボ.155pp. 神戸市スポーツ教育公社,神戸.
Ando, H., 1962. The comparative embryology of Odonata with special reference to a relic dragonfly Epiophelebia superstes Selys. The Japan Society for the Promotion of Science, Tokyo.
安藤 裕,1996.トンボ目 Odonata.安藤 裕・小林幸正監修,昆虫発生学 上.培風館,東京.
新井 裕,2001.トンボの不思議.165pp., どうぶつ社,東京.
Becnel, J.J. and Dunkle, S.W., 1990. Evolution of microphytes in dragonfly eggs (Aisoptera). Odonatologica 19:235-241.
*Bennett, S., and & Mill, P.J., 1995. Lifetime egg production and egg mortality in the damselfly Pyrrhosoma nymphula (Sulzer) (Zugoptera: Coenagrionidae). Hydrobiologia 310:71-78. (in Corbet, 1999).
*Brandt, A., 1869."Beiträge zur Entwicklungsgeschichte der Libelluliden und Hemipteren." Mém. Acad. St Pétersbourg (Petrograd), Ser. 7, Tome XIII, pp.1-33, 3plates. (in Tillyard, 1917).
Corbet, P. S., 1999. "Dragonflies Behavior and Ecology of Odonata". Cornell Universitiy Press. New York.
井上 清・尾花 茂・冨士久芳久,1981.アオヤンマの生活史.Tombo 23(1/4):23-27.
神宮字寛・露﨑 浩・佐藤照男,2006.アキアカネ卵の孵化におよぼす光と水温の関係.Trans. of JSIDRE, 243:79-84 (2006.6).
桐谷圭治,1997.日本産昆虫,ダニ,線虫の発育零点と有効積算温度.農業環境技術研究所資料 21:1-72.
*Laplante, J.-P., 1975. Observations sur la ponte de quatre odonates du genre Lestes (Zygoptera: Lestidae) au Quebec. Nat. Can. 102:279-292. (in Corbet, 1999).
*Miall, L.C., 1985. "The Natural History of Aquatic Insects". (Dragonflies, chapter IX, 328-345. (in Tillyard, 1917).
*Pierre, A., 1904. L'éclosion des œufs du Lestes viridis Van der Lind.. Ann. Soc. Ent. France, LXXIII, pp. 477-484, 1 plate. (in Tillyard, 1917).
*Pilon, J.G., and Masseau, M.J., 1984. The effect of temperature on egg development in Zygoptera: a preliminary discussion. Advances in Odonatology 2:177-193. (in Corbet, 1999).
齊藤四海智・先崎悠介・米澤千夏・千葉克己・神宮字寛,2017.卵と幼虫の発育ゼロ点と有効積算温度を用いたアキアカネ保全に有効な中干し実施日の検討.IDRE Journal 304: I_37-I_46 (2017.6).
新村捷介,2001.アカネ属の産卵方法と卵の形との関係.Gracile (63):1-7.
新村捷介,2004.トンボ卵に寄生する卵寄生蜂.Gracile (67):1-8.
新村捷介,http://tombon.com/EGG2.htm.近畿のトンボ雑記.2026.6.4 閲覧.
曽根原今人,1982.八ヶ岳 オオトラフトンボの生活史.203pp., 社団法人 信濃教育会出版部,長野.
Tillyard,R.J., 1917. "The biology of dragonflies". Cambridge Univ. Press, Cambridge.
上田哲行,1993.山へ上るアキアカネ,上らないアキアカネ アキアカネの生活史における諸問題-1.インセクタリウム 30:292-299.
渡辺庸子,2010.トンボの発生学.日本環境動物昆虫学会編 改訂 トンボの調べ方:106-118.
吉田雅澄,2014.飼育による日本産トンボの卵・幼虫期.Aeschna (50):1-12.