嘥丏奺搰偺柺愊偲偦偙偵暘晍偡傞僩儞儃偺庬悢偺娭學
丂偝偰丆偙偆偄偭偨抧巎傪帩偮撿惣彅搰偺搰乆偵偼丆偳偺傛偆側摿挜傪帩偭偨僩儞儃偨偪偑惗懅偟偰偄傞偺偱偟傚偆偐丏傑偢嵟弶偵丆奺搰乆偵惗懅偡傞僩儞儃偺庬悢傪斾傋偰傒傑偟傚偆丏惗懺妛偱傛偔抦傜傟偨娭學偵丆乽柺愊庬悢嬋慄乿偲偄偆偺偑偁傝傑偡丏搰偵惗懅偡傞惗暔偺庬悢偲搰偺柺愊偵偼憡娭娭學偑偁傞偲偄偆傕偺偱偡丏偦偙偱丆撿惣彅搰偺僩儞儃偵偮偄偰挷傋偰傒傑偟偨丏
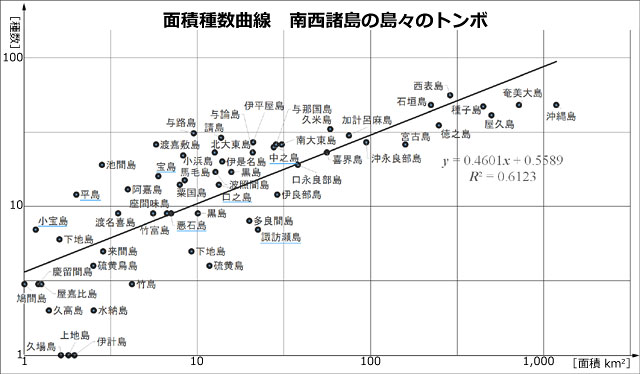
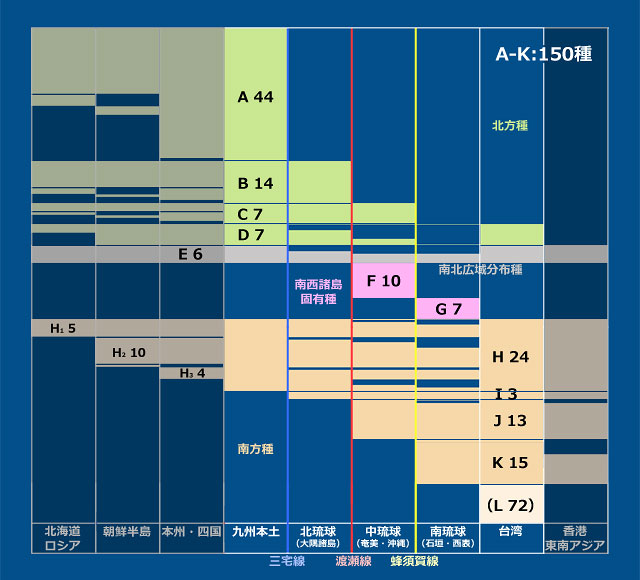
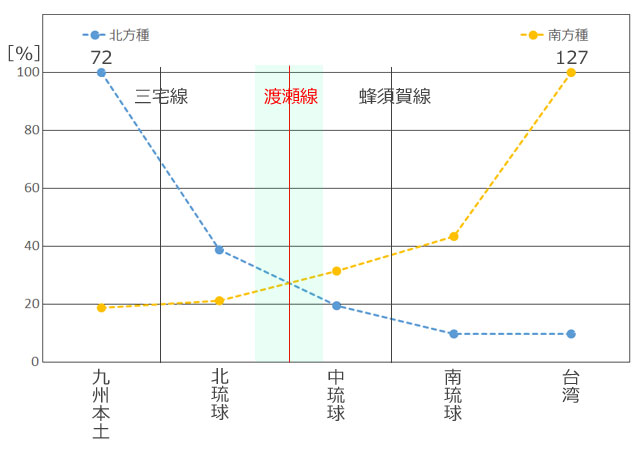
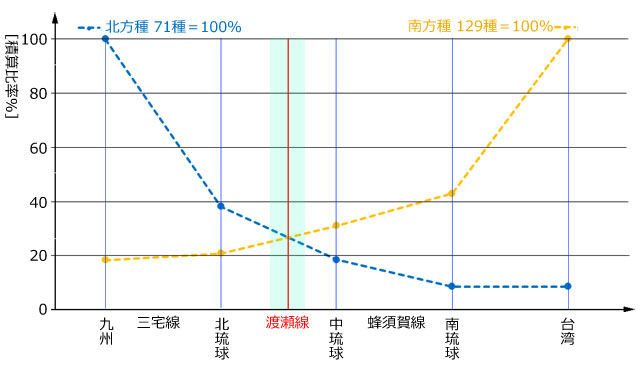
丂僩儞儃暘晍偺僨乕僞偼峕暯(2023)丆旜墍傜(2007)偺暘晍昞(愲妕彅搰偼彍偔)丆旜墍傜(2022)偵丆偦偺屻偺抦尒(戜榩傪彍偔)傪壛偊偨傕偺傪梡偄傑偟偨丏柺愊庬悢嬋慄偺応崌乽偦偺搰偵惗懅偡傞庬悢乿傪埖偄傑偡偺偱丆嬼嶻庬偼彍偒丆傑偨垷庬傪庬偲偟偰傑偲傔傞偲丆偪傚偆偳100庬偵側傝傑偟偨丏奺搰偺柺愊偼栚嶈(1985)偵宖嵹偝傟偰偄傞抣傪梡偄傑偟偨丏偦偟偰昤偄偨偺偑恾4偱偡丏憡娭學悢偼0.78偲丆傑偢傑偢偺寢壥偑弌偰偄傑偡丏側偍 Karbe et al.(2025) 偵傛偭偰屌桳庬傾儅儈僆僯儎儞儅偲屌桳垷庬儎儞僶儖僆僯儎儞儅偑婰嵹偝傟傑偟偨偑丆幏昅帪婜偺娭學偐傜昞侾埲奜偺暘愅偵偼斀塮偝傟偰偄側偄偙偲傪偍抐傝偟偰偍偒傑偡丏
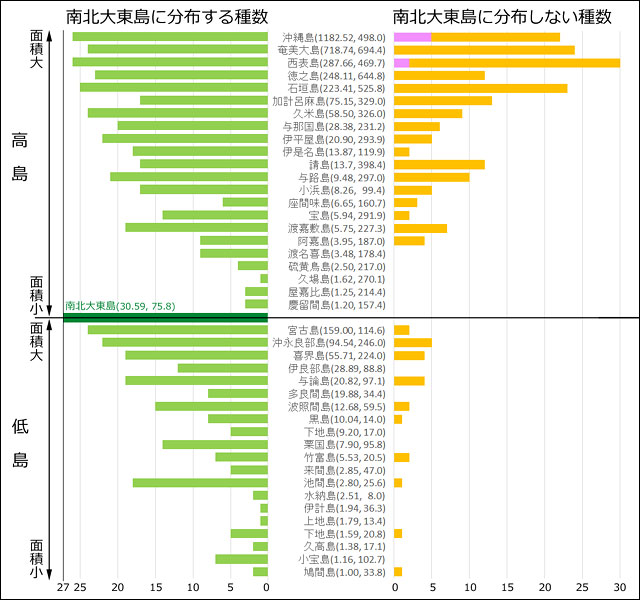
恾5丏撿惣彅搰(愲妕彅搰傪彍偔)偺僩儞儃偺柺愊庬悢嬋慄丏庬悢丆柺愊偼忢梡懳悢曄姺傪偟偰偄傞丏憡娭學悢偼0.78丏堦斒偵丆柺愊偺戝偒偄搰偵傛傝懡偔偺庬偑惗懅偟偰偄傞偲偄偊傞丏栚嶈(1985)丆旜墍傜(2007)丆峕暯(2023)丆偦偺懠偺忣曬傪傕偲偵昤夋丏惵壓慄偼僩僇儔楍搰偺搰乆丏
丂偙傟偼丆柺愊偑戝偒偗傟偽丆偝傑偞傑側惗懅娐嫬偑搰撪偵懚嵼偡傞壜擻惈偑崅傑傝丆娐嫬梫媮惈偺堎側傞傛傝懡偔偺惗暔庬傪彽偒擖傟傞偙偲偑壜擻偵側傞偨傔丆偲峫偊傜傟偰偄傑偡丏僩儞儃偵娭偟偰偙偺偙偲傪偍榖偟偟偰偍偒傑偟傚偆丏
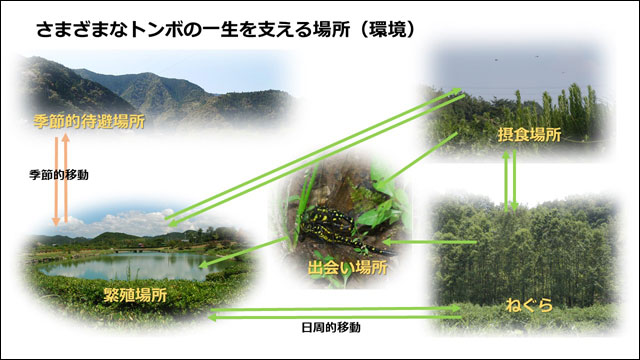
丂僩儞儃偺惗懅傪壜擻偵偡傞偵偼丆婥岓側偳偺暔棟揑娐嫬偑岲揔偱偁傞偙偲埲奜偵傕丆惗妶巎慡斒傪巟偊傞応強偑昁梫偱偡丏恾5偼僩儞儃偺堦惗傪巟偊傞偺偵昁梫側応強傪帵偟偰偄傑偡丏偙傟傜屄乆偺娐嫬偵懳偡傞梫媮惈偼丆僩儞儃偺庬椶偵傛偭偰彮偟偢偮堎側偭偰偄傑偡丏斏怋応強傪椺偵偲傞偲丆悈堟偺惈幙(幖抧丆壨愳丆抮徖側偳)傗悈堟偍傛傃傑傢傝偺怉惗偺梫媮惈偑庬偵傛偭偰偐側傝僔價傾偵堎側偭偰偄傑偡丏偟偨偑偭偰丆懡條側悈曈娐嫬偑懚嵼偡傟偽丆傛傝懡偔偺娐嫬梫媮惈傪帩偮僩儞儃庬偑偦偙偱斏怋偱偒傞偙偲偵側傝傑偡丏偦偺偨傔偵偼丆傗偼傝偦傟側傝偺搰偺柺愊偑昁梫偵側偭偰偔傞偱偟傚偆丏
恾6丏僩儞儃偺惗妶巎偵昁梫側応強(娐嫬)偺奣擮恾丏偙傟傜偼偁偔傑偱奣擮揑偵暘椶偟偨傕偺偱丆嬻娫揑偵偼廳暋偟偰偄傞応崌偑懡偄丏椺偊偽丆懡偔偺僩儞儃偱偼弌夛偄応強偲斏怋応強偑摨堦偱偁傞丏撿惣彅搰偺僩儞儃偵偼丆偍偦傜偔傾僉傾僇僱偺傛偆側婫愡揑懸旔傪峴偆僩儞儃偼偄側偄偲巚傢傟傞丏
嘦丏搰乆偺僩儞儃憡偺摿挜
丂崱傑偱偺専摙寢壥偐傜丆奺搰乆偵惗懅偡傞僩儞儃偵塭嬁傪梌偊傞庡側梫場偲偟偰丆婥岓忦審丆抧巎丆抧棟揑妘棧偺掱搙丆搰偺柺愊丆惉場偲悈暥宯偵娭學偡傞崅搰丒掅搰偺嬫暿丆憡懳揑側奀柺偺忋壓摦丆僩儞儃偺堏摦擻椡側偳偑嫇偘傜傟傞偱偟傚偆丏偙傟傜偼偦傟偧傟撈棫側傕偺偱偼側偔丆屳偄偵娭學偟崌偭偰偄傞傕偺偱偡丏巹偑朘傟偨偄偔偮偐偺搰乆偵偮偄偰偼暿崁偱弎傋傞偲偟偰丆偙偙偱偼丆僩儞儃偺堏摦擻椡偲奺搰乆偺僩儞儃憡丆屌桳庬偺暘晍忬嫷偵偮偄偰丆慡懱揑偵挱傔偰傒偨偄偲巚偄傑偡丏
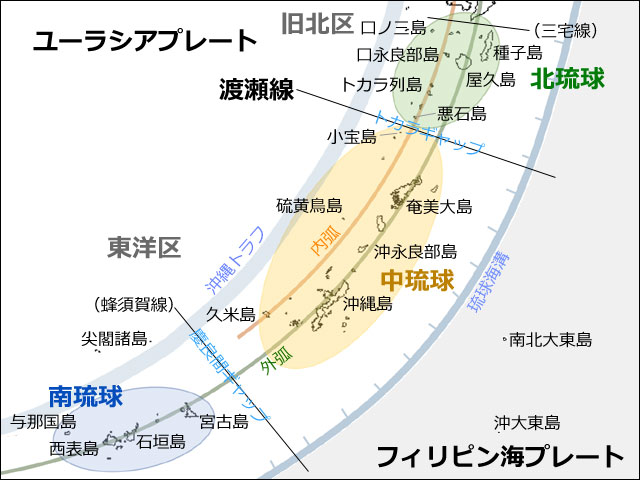
丂撿惣彅搰偼棧搰偺廤崌懱偱偡偐傜丆僩儞儃偺暘晍偲奀傪搉傞擻椡偵偼枾愙側娭楢偑偁傞偼偢偱偡丏偦偙偱丆搉奀壜擻側僩儞儃偑奺搰乆偵偳傟偔傜偄偄傞偐傪挷傋偰傒傞偙偲偵偟傑偟偨丏偨偩偙偺偲偒丆偁傞僩儞儃偑奀傪搉傞擻椡傪桳偡傞偐偳偆偐傪丆乽媞娤揑偵乿偳偆傗偭偰敾掕偡傞偐偑旕忢偵擄偟偄栤戣偵側傝傑偡丏偙偆偄偭偨偲偒偵桳岠側偺偑憖嶌揑偵掕媊偡傞曽朄偱偡丏
丂偦偙偱丆堦搙傕棨懕偒偵側偭偨偙偲偺側偄奀梞搰偱偁傞撿杒戝搶搰偵拲栚偟傑偟偨丏壂撽杮搰偐傜搶偵360km傎偳棧傟偨偙偺搰偺僩儞儃偼丆偡傋偰奀傪墇偊偰旘棃偟偨偲峫偊傞偺偑崌棟揑偱偡丏偦偙偱彮側偔偲傕撿杒戝搶搰偱婰榐偝傟偨僩儞儃偼奀傪搉傞擻椡偑偁傞偲掕媊偡傞偺偱偡丏偙偺傛偆偵丆崌棟揑側暿偺娤應傗帠幚偐傜掕媊偡傞曽朄傪憖嶌揑掕媊偲偄偄傑偡丏偨偩偟堦偮拲堄偡傞傋偒偙偲偼丆偙偺掕媊偱偼丆撿杒戝搶搰偵暘晍偟側偄僩儞儃偵堏摦椡偑側偄偲偼偄偊側偄偙偲偱偡丏
丂偝偰丆傑偢柺愊庬悢嬋慄傪昤偄偨帪偵巊偭偨僨乕僞傪梡偄偰丆奺搰偵惗懅偡傞僩儞儃傪丆撿杒戝搶搰偵暘晍偡傞庬偲偟側偄庬偵暘偗傑偟偨丏偨偩撿杒戝搶搰偼搉悾慄偺撿懁偵埵抲偡傞偺偱丆崱夞偼搉悾慄偺撿懁偵偁傞搰偩偗傪懳徾偲偟傑偟偨丏傕偟偙傟傪棶媴屖慡懱偵峀偘傞偲丆椺偊偽丆戝嬿彅搰(庬巕搰傗壆媣搰)偵暘晍偡傞恾7偺僒僽僌儖乕僾B偺僩儞儃偨偪偼撿杒戝搶搰偵偼慡偔尒傜傟傑偣傫丏偙傟傜傪丆堏摦椡偺掕媊偲偟偨撿杒戝搶搰偺僩儞儃憡偲斾妑偡傞偺偼揔愗偱偼側偄偲峫偊偨偐傜偱偡丏
丂偝偰埲忋偺傛偆側忦審壓偱昤偄偨僌儔僼偑丆恾7偱偡丏
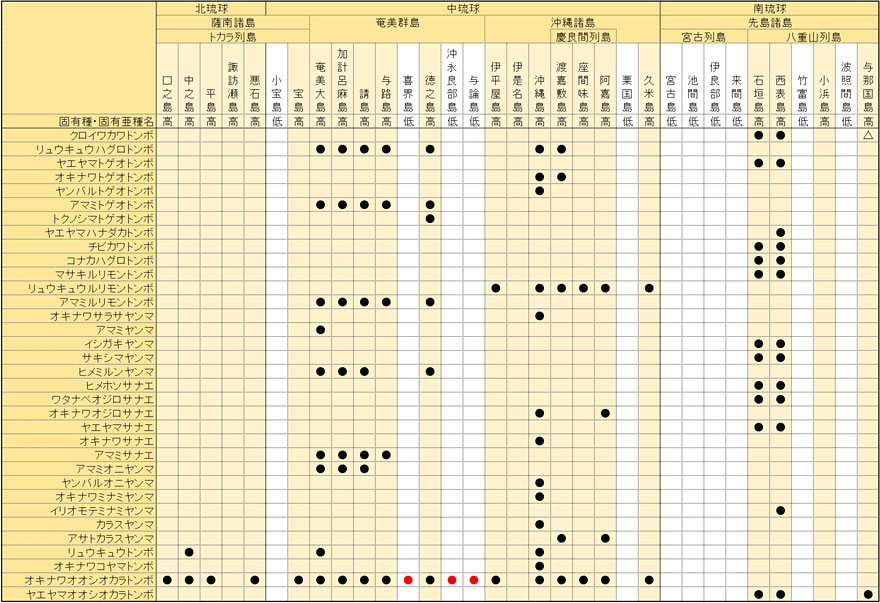
恾7丏搉悾慄埲撿偺奺搰乆偺僩儞儃憡傪撿杒戝搶搰偺偦傟偲斾妑偟偨傕偺丏崅搰偲掅搰偵暘偗丆偝傜偵偦傟偧傟傪柺愊偺崀弴偱昞帵偟偨丏(丂)撪偺悢帤偼偦傟偧傟(柺愊[km
2]丆嵟崅昗崅[m])偱偁傞丏撿杒戝搶搰偵偼27庬偑暘晍偡傞丏僺儞僋怓偼偦偺搰偵偟偐偄側偄屌桳庬偱偁傞丏旜墍傜(2022)偵傛傞偲丆
撿戝搶搰偵暘晍昞帵偑偁傞僩儞儃偼28庬偁傞偑丆偦偺偆偪僗僫傾僇僱傪彍偄偰27庬偲偟偰偄傞丏
丂埲壓乽撿杒戝搶搰偵暘晍偡傞庬乿偼乽堏摦椡偺偁傞庬乿丆乽撿杒戝搶搰偵暘晍偟側偄庬乿偼昁偢偟傕堏摦椡偑側偄偲偼偄偊側偄偺偱偡偑丆偙偙偱偼棟夝偟傗偡偔偡傞偨傔乽堏摦椡偑側偄偲峫偊傜傟傞庬乿偲尵偄姺偊傞偙偲偵偟傑偡丏
丂傑偢偙偺僌儔僼偵傕丆柺愊亅庬悢偺娭學偑怓擹偔昞傟偰偄傞偙偲偑暘偐傝傑偡丏偦偟偰柺愊偑彫偝偄搰(栺3.5km
2埲壓)偵暘晍偡傞僩儞儃偼丆崅搰丒掅搰偵偐偐傢傜偢丆傎偲傫偳偑乽堏摦椡偺偁傞庬乿偱愯傔傜傟偰偄傑偡丏傑偨掅搰偵暘晍偡傞僩儞儃偼丆柺愊偺戝彫偵偐偐傢傜偢丆乽堏摦椡偑側偄偲峫偊傜傟傞庬乿偑彮側偔乽堏摦椡偺偁傞庬乿偑懡偄孹岦偑尒傜傟傑偡丏椺偊偽丆掅搰偱傕斾妑揑柺愊偺戝偒偄媨屆搰傗壂塱椙晹搰偼丆崅搰偺暲傃偱偼愇奯搰偲壛寁楥杻搰偺娫偵擖傝傑偡偑丆乽堏摦椡偺偁傞庬乿偺悢偼偁傑傝曄傢傜側偄偺偵丆乽堏摦椡偑側偄偲峫偊傜傟傞庬乿偼彮側偔側偭偰偄傑偡丏
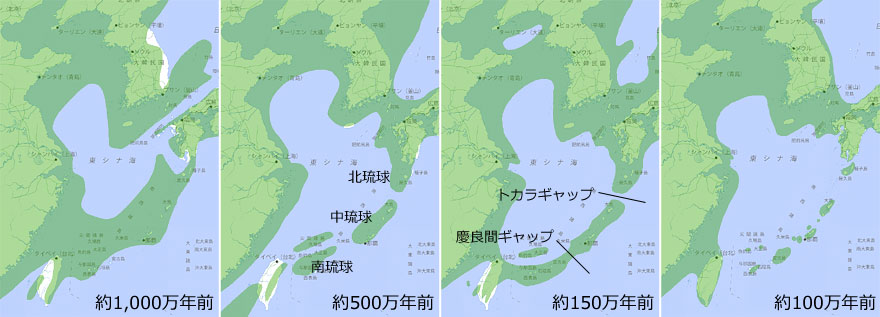
丂撿惣彅搰偼丆夁嫀偺抧妅曄摦傗奀柺曄摦偵傛偭偰丆憡懳揑側奀柺偺埵抲曄壔偵偝傜偝傟偰偒偨偼偢偱偡丏偦傫側拞偱丆嵟崅昗崅偺崅偄崅搰偼柺愊傕峀偔側傞孹岦偑偁傝丆夁嫀偵奀柺忋徃偟偨帪戙偵傕嶳抧丒怷椦傗壨愳宯偑偦傟側傝偺峀偝偱堐帩偝傟偰偄偨偲峫偊傜傟傑偡丏偙偆偄偭偨娐嫬偺傕偲偱丆堏摦惈偺朢偟偄庬傕尭彮傑偨偼愨柵偡傞偙偲側偔巆懚偡傞偙偲偑偱偒偨偺偱偟傚偆丏偝傜偵偙偺妘棧偝傟埨掕偟偨娐嫬偺拞偱丆屌桳庬偝偊恑壔偡傞偙偲偑偱偒偨偲峫偊傜傟傑偡丏