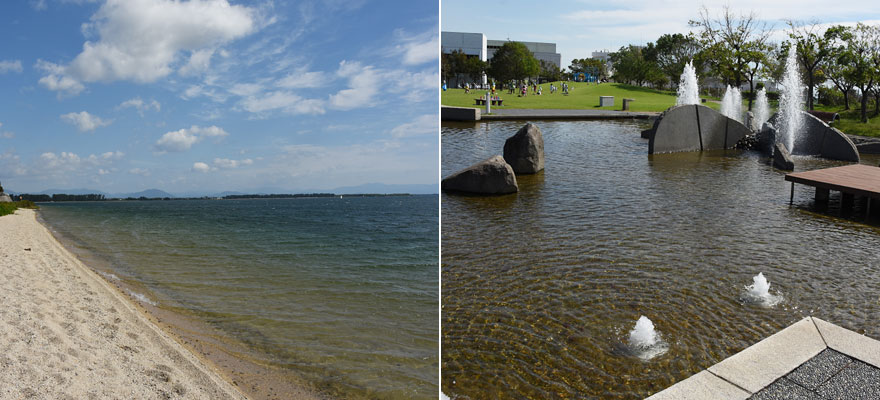トンボの生態学
2.生息地選択 目次
トンボの生態学 2.生息地選択
トンボは自ら生息地を選択しているのか?
トンボの生息地選択という問題を考える場合,トンボに「自ら『新規に』生息地を選択する」という事実があるかどうかを,まず検討しておく必要があります.その際,空を飛翔し自由に移動できる成虫のステージで生息地を選択していることは,ほとんど疑いのない事実としてよいでしょう.ただ幼虫でも,オジロサナエのように,河川をかなりの距離流下移動する種が知られています(新井,1989).新井(1993)はこの流下が増水による受動的なものである可能性が低いことを指摘していて,自ら生息地を選択している可能性があるかもしれません.この節では,このような事例は扱わず,成虫の生息地選択を中心にしてお話を進めていきます.
トンボの成虫は,幼虫時代を過ごした水域で羽化すると,通常いったん水辺を離れます.離れる距離はトンボの種類によってさまざまです.例えば,オオキトンボは,6月ころに平地のため池などで羽化した後,未熟成虫は一気に空高く舞い上がり,その池を離れます.その際かなり遠方に飛散しているようで,しばらく羽化した池の周辺では成虫を目にすることはまずありません.9月下旬の繁殖時期になると,どこからともなくその池にやって来て,交尾や産卵活動を行います.オオキトンボが好きな産卵環境は,水落をしてなだらかな泥の水際が出現しているような場所です.このとき,この池が水落をしていなかったら,やって来る個体数は非常に少ないか,または 0 のときもあります.逆に産卵に適した状態であれば,たくさんの個体が集まってきます.このような個体数変動がある事実は,とりもなおさず,池の状態を成虫が判断している結果だと考えられます.
写真1.オオキトンボは羽化すると空高く飛んで一気にその池を離れる(左).秋になると,水落されてなだらかな泥の水際ができる池に集結する(右).
一方ベッコウトンボは羽化水域からそれほど遠くへ離れないトンボであるとされています(例えば浜田・井上,1985;杉村ら,1999;山本ら,2009).三時・平田(1997)のマーキングによる飛散調査では,1km以遠での確認頻度は非常に低くなると記しています.私が神戸市内の生息地で実際に調査した結果(青木,1997)では,未熟成虫は,唯一の繁殖池が存在する谷筋約700mの範囲にのみ分散していました.分散域内には池が3つありますが,成熟オスが繁殖活動を見せたのは,羽化した池の他に,ベッコウトンボが好むようなヨシの茂った,2つの池の一部の範囲でした.羽化した池以外にもオスが集結した事実は,帰巣性などの能力によるものではなく,オスがその場所を自ら選択した結果だと判断できます.このように,羽化水域を大きく離れないトンボも,新たに繁殖活動を行う場所の選択を行っています.
ベッコウトンボでも,羽化した池を自ら離れ,移住してしまうことがあります.兵庫県下では,多くのトンボ研究家が熱心に探索したにもかかわらず,2001年時点でベッコウトンボの生息している池は,互いに近接した2カ所だけしか知られていませんでした.その2001年に私は,そこから直線距離で約10km離れた2つの異なる場所で,オスのベッコウトンボを目撃しました*1.もちろんその池やその近くを調べても,他のベッコウトンボの姿はありませんでした.産地数が著しく減少したベッコウトンボの,こういった既産地から大きく離れた場所での発見は,これらの個体がもとの産地から移住した結果だという可能性が高いでしょう.静岡県でも未熟なメスが既産地から17km離れた場所で発見されています(三時・平田,1997).これらの事実は,毎年同じ生息場所で繁殖を繰り返している種でも,「自ら」移住して新しい生息場所を探す行動をとることを示しているといえます.
以上よりもっと確かな例として,新しいトンボ池を造成すると,間もなくオツネントンボやクロスジギンヤンマなど,何種類かのトンボがやって来ます.これ一つをとっても,トンボは「自ら『新規に』生息地を選択する」ことができるといえます.
以上の例から,トンボ成虫が,生息地を自ら探し選択できる能力を有していることは,間違いない事実といえます.
写真2.人工的に造成されたビオトープ池(トンボ池)(左).トンボ池で羽化したマルタンヤンマのオス(右),ただし左の写真とは異なる池である.
トンボの生態学 2.生息地選択
トンボの生息場所
トンボは実にさまざまの陸水環境に適応放散しています.陸水は大きく分けて流水と止水に分類されます.
Ⅰ.流水
流水である河川は,可児(1944)によれば,上流,中流,下流に分けられます.これは感覚的な分類ではなく,河川形態に注目した分類です.自然の河川は,上,中,下流を問わず,一般に蛇行して流れています.可児は,この一つの蛇行区間における瀬と淵の数に注目して,まず上流と中・下流に分けました.上流というのは,一つの蛇行区間内に,複数の瀬と淵が存在し,瀬から淵へ入るときに段差があるような流れを指します(Aa型).中流と下流には,一つの蛇行区間に瀬と淵がそれぞれ一つずつしかありません(B型).そして,瀬から淵への流れ込む水が波立っている場合(Bb型)を中流,波立たない場合(Bc型)を下流と分けました.
河川間で得られた生物の調査結果を比較するためには,この分類は非常に有用です(水野・御勢,1993).しかしトンボの生息環境を議論するときには,やや使いづらいと思っています.例えば,同じ上流でも,河川の幅や水量がずいぶん違う場合があり,この分類ではそれらは特に区別されません.日当たりがよく幅広くて水量の多い上流と,樹林におおわれ薄暗くて細く流量も少ない上流では,生息するトンボは微妙に違ってきます.これは,トンボの成虫による生息地選択には,陸上の環境が大きく影響していると考えられるからです.さらに,現在多くの河川は,護岸工事,浚渫,堰の造成などの治水・利水工事が行われています.その結果蛇行や瀬・淵のある自然の河川は少なくなってきましたし,堰にせき止められた流れの緩い部分に砂が分厚く堆積したような環境も,上・中・下流を問わず現出しています.さらにまた潅漑用水路や疎水のような人工的流水,湿地の中の細流といった,河川の分類に収まりきらない流水環境もあります.トンボたちはそういった環境に入り込んで生活しています.
以上からここでは,可児(1944)の分類したAa型の河川とそこに流れ込む支流からなる流域とその周辺の陸上環境をまとめて上流域,Bb型およびBc型,あるいは治水・利水工事がなされていても自然な状態ならBb型およびBc型になりそうな河川とその周辺の陸上環境をまとめて,それぞれ中流域および下流域と呼ぶことにします.さらに,上流域で,湧水近くの流量があまり多くない比較的勾配のある流れを「源流」,湿地や山裾の湧き水に端を発し勾配が緩い細い流れを「細流」,下流域で,河口近くの海や湖の影響を受けている流域を「河口域」,また人工的な流水を用途に応じて「溝川」や「用水路」や「疎水」などと呼ぶことにします.多くのトンボたちは,これら多様な流水域・流水環境に入り込んで生活しています.

写真3.上流.左:源流,湧水のすぐ下の上流.右:上流(Aa型).瀬と淵が一つの蛇行区間(青矢印)に複数存在し,瀬から段差を持って淵に入る.
源流にはムカシトンボ,クロサナエ,ヒメクロサナエ,アサヒナカワトンボ,ミナミヤンマなどが見られる.
Aa型の上流には,源流に見られるトンボ以外に,オジロサナエ,ダビドサナエ,ミヤマカワトンボ,ミルンヤンマなどが見られる.
写真4.中流(Bb型).左:上流に近い.右:下流に近い.瀬と淵が一つの蛇行区間(青矢印)に一つだけ存在し,淵に入る瀬に波が立っている(白矢印).
上流に近いBb型の中流は,サナエトンボ科,カワトンボ科,ヤマトンボ科,コシボソヤンマなど,もっとも多種のトンボが見られる流水域である.
下流に近いBb型の中流になると,見られるトンボの種類は急激に少なくなり,川によってはミヤマサナエなどのトンボが見られる.
写真5.下流(Bc型).水底には瀬や淵があるのかもしれないが,表面的にはほとんど平坦な水面である.これは河口近くなので写真は河口域でもある.
メガネサナエ属のトンボが見られる.流れが緩いのでギンヤンマ,コフキトンボ,オオヤマトンボのような止水性の種も現れる.
写真6.左:細流.湿地の中を流れる砂泥底の浅い流水.右:用水路.潅漑用の素掘りの用水路.
こういった流水には,オニヤンマ,キイロサナエ,ヤマサナエをはじめ,止水性のオオシオカラトンボなども見られる.
Ⅱ.止水
トンボの生息場所から見た止水にも,さまざまのものがあります.まず自然の池というのは,私たちの身の回りにはもうほとんどなくなってしまいました.身近な池といえば潅漑用のため池です.さらに河川にできるワンドや湿地の小さな水たまりも止水といえます.大きなものとしては湖があります.大きな湖は他の止水と違い,流水性のトンボを育むことができます.新しく人工的につくられたものとしては,ダム湖,調整池,公園の噴水池,ビオトープ池などがあります.トンボたちはこういった止水に入り込んで生活しています.
トンボが生息している止水の中には,非常に特殊なものが存在します.それはファイトテルマータと呼ばれる止水環境です.具体的には,柵に使っている竹の先を切った部分の水たまり,葉腋と呼ばれる葉のつけ根部分にたまった水,あるいは樹洞の中にたまった水のような場所です.こういった場所に生息するトンボは主に熱帯地方で見られます.日本産のトンボでは,場所は台湾ですが,キイロハラビロトンボの幼虫がファイトテルマータにいたのが見つかっています(連・松木,1979).
写真7.自然の池.
左:高地には,溶岩でせき止められてできた自然の池が残っている.信州では,ルリイトトンボなどの寒冷地のトンボが見られる.
右:高地にできた池が遷移し高層湿原となり,こういった沼が生じる.信州では,ムツアカネ,カオジロトンボ,エゾイトトンボなどが見られる.
写真8.潅漑用ため池.植生豊かな池ではいずれも多種のトンボが見られるが,汚染されたり,管理されすぎるとトンボが少なくなる.
左:平地に堰堤を築いてつくられた皿池.低い位置にあり,雨水などで養分が流れ込むので,富栄養で植生豊かである.
右:谷の下手に堰堤を築き湧水などをせき止めてつくられた谷池.高い位置につくられるので養分の流れ込みが少なく,貧栄養であることが多い.
写真9.左:湖.湖岸に波が打ち寄せ,溶存酸素量が多くなるためか,流水性の種が生息している.
右:都市公園の噴水池.こういったところにも,ウスバキトンボ,タイリクアカネ,コノシメトンボなどが産卵にやって来る.
Ⅲ.流水性種と止水性種
さて,陸水環境の流水と止水について簡単に述べました.実は,トンボには流水環境に生息する種*1と,止水環境に生息する種*2とがあって,前者を流水性種,後者を止水性種とよんでいます.トンボはかなりはっきりとこれらに二分されます(Corbet, 1999).この違いに系統性はあまり関係ないようで,分布拡大していったそれぞれの環境で,種ごとに独自に獲得した形質のようです.というのは,同種の間でも,地域によって,流水に好んで入り込む個体群と,止水で主に見られる個体群が存在するという,行動の分化が見られるからです.例えばベニトンボは,沖永良部島では多くが流水環境に入り込んでいますが,北限近くの徳島県などでは主に池で見いだされます.
流水性種と止水性種に二分される理由は,それぞれの種の幼虫で溶存酸素に対する要求量に違いがあるからと考えられています(Wright & Shoup, 1954 in Corbet, 1999).流水環境は,水の動きによって酸素が溶け込む機会が多くなり,一般に止水に比べて溶存酸素量が高くなる傾向にあります.溶存酸素が幼虫生存の制限要因になっていることは,止水性種の幼虫は流水環境で見つかることはしばしばあるがその逆はあまりない,という事実から十分予想できます.湖に流水性種が入り込める理由は,波打ち際の水の動きが溶存酸素量を高めるからだと考えられます(Corbet, 1999).またベニトンボのような例では,南方では気温が高くなり,止水の溶存酸素量が減少するため,より溶存酸素量の高い流水環境に入り込むのだと解釈できます.
この事実は,生息地選択をする成虫に,重大な判断を迫ることになるはずです.流水性種は間違って止水環境に産卵してしまうと,幼虫の生存率が下がって,次世代の個体数に大きな影響を与えるかもしれないからです.したがって,特に流水性種は,流水を止水と区別する能力を,その進化の過程で獲得していると考えねばなりません.
*1.流水性種 lotic species
*2.止水性種 lentic species
写真10.流水性種と止水性種.左:流水性種のコヤマトンボメスの羽化,右:止水性種のオツネントンボオスの羽化.
トンボの生態学 2.生息地選択
トンボの視覚と生息地選択
長い間トンボの観察を続けていると,例えば池とその周辺の環境を見ただけで,多分この種がいるはずという見当が付くようになります.このとき,その水域の植生の状態,水の状態,周辺の樹林の状態等々を,私たちは「目」で判断しているわけです.このことはとりもなおさず,視覚情報によって,生息地の適切な判断が可能なことを示しています.
では,トンボはどうでしょう.トンボも視覚情報を使って生息地を選択しているのでしょうか.トンボには大きな複眼が備わっています.この複眼で何を見ているのか,ここでトンボの視覚世界について,少し触れておきましょう.
Ⅰ.トンボの視覚世界
Futahashi et al.,(2015)は,16種類の日本産のトンボを使って,複眼の視細胞のオプシン(色に感じる視物質)遺伝子の発現状態を調べました.ヒトは赤・緑・青を感じる3種類のオプシンを網膜の錐体細胞に持っています
*1.ところが,トンボの場合,なんとオプシンの遺伝子は15-33もある(つまりオプシンタンパク質も同じ数あるといえる)そうです.この数からだけでも,トンボは非常にカラフルな視覚世界を持っていると考えられます.これらのオプシンは,それが感じることのできる波長によって,紫外線領域,短波長領域,長波長領域のものに分類されています
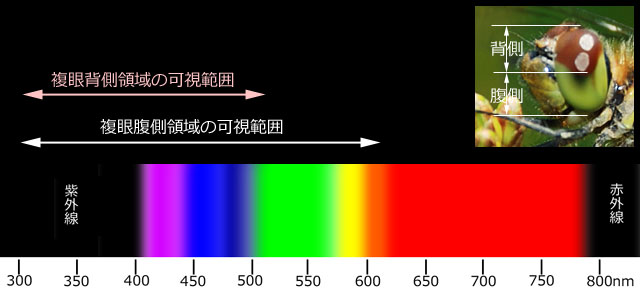
*2.そして,これらの遺伝子は,複眼の背側と腹側で,その発現状態が違うことが分かりました.アキアカネでは,複眼背側では短波長領域の,複眼腹側では長波長領域の遺伝子が,それぞれより多く発現していました.紫外線領域のオプシン遺伝子はどちらの領域でも発現していました.そしてこの発現傾向は,他の不均翅亜目でも同様でした.ただし均翅亜目では少し異なっているようです.アキアカネ複眼の背側・腹側それぞれの波長感受性範囲を図1に示しました.
図1.アキアカネのオプシンの感受性範囲 (Futahashi et al., 2015より作成).
アキアカネのオプシンの数は,紫外線領域1,短波長領域5,長波長領域10である.
少し話が脱線しますが,この可視領域を見て気づくのは,アキアカネは赤の領域を感受していないということです.つまり,アカトンボの赤い色がほとんど見えていなくて,黒く見えると予想されます(写真11左).
さて,この可視範囲から次のようなことが考えられます.トンボは空中を飛ぶ動物です.そのとき複眼背側は空の方を向いています.日中の空全体から来る光には短波長(青い色)の光が多く含まれています.だから晴れた空は青いわけですが,これは,複眼背側が主に青い光を感じることと一致しています.トンボの大敵は鳥ですし,トンボの餌は空中を飛ぶ小昆虫です.これらが青い空の影になったとき,そのコントラストから発見が容易になるでしょう.また下方および側面は緑や黄色の長波長の光をよりよく感じています.茶色い土や緑の植生に反射した光は長波長のものが多いので,それを感じることができる複眼腹側は,下方の地形などの詳細な状況を判断するのに有効に働いていると考えられます.
さらにトンボは,水面を見るときに偏光を感じているのではないかとも考えられていて(Schwind, 1991 in Corbet, 1999),もしそうであるならば,偏光を多く含む水面の反射光を効率的に見分けることも可能であると考えられます.
以上を生息地選択の文脈でまとめますと,複眼背側の視覚は,生息地に日向や日陰がどのように分布するかを判断するのに役立つでしょうし,複眼腹側の視覚は,生息地の植生状態,水の状態,水際の状態などを判断するのに役立ちます.つまり複眼は,生息地選択に使うデバイスとして,十分な能力を有しているといってよいでしょう.
*1.三原色の混合でヒトが感じることのできる全ての色をつくることができるというのは,このオプシンの数に対応しています.
*2.オプシン遺伝子の数は,紫外線領域のものは1個ですが,他は複数のオプシンが含まれます.
写真11.左:赤い色成分を消し去ったアキアカネの画像.右:トンボの複眼背側と腹側の色覚の違いが持つ意味を示した模式図.
左:トンボにはこのように見えているのだろうか? もっとも紫外線の影響を考えていないので,もっと違う色覚を生じているかもしれない.
Ⅱ.トンボの視覚と生息地選択
では,トンボの視覚が,本当に生息地選択に役立っているという証拠はあるのでしょうか.これについては,海外の研究者が,いくつかの実験的研究を行っています.
Wildermuth & Spinner (1991) は,オーストリアのチロル地方で,ホソミモリトンボ Somatochlora arctica を用いて選択実験*3を行いました.自然の産卵場所(2m2),および黒くて光を反射させるプラスチックホイル(2×4mと1×1m),光を反射しない木綿の黒布(1×1m),および黒の絹布(1×1m)などのダミーを,地面をはじめとしたさまざまの背景の上20cmの位置に伸ばして敷いて,そこにトンボが引き寄せられるかということを定量的に調べました.その結果,オスもメスも光を反射するプラスチックの上で,ホバリングしたり,探索飛翔したり,あるメスは産卵行動さえとりました.これは「光っている」ということを視覚によって認知しているからだと考えられます.
Wildermuth (1993) は,上と類似の実験を,ルリボシヤンマを使って行いました.やはり黒くて光を反射するプラスチックホイルにオスメスとも引き寄せられ,繁殖行動の全てのレパートリーがその上で行われました.この実験では白い色の木綿や絹の布もダミーとして用いられていましたが,これらにはほとんど引き寄せられませんでした.結論として,黒い色のバックに光が反射している場所を,産卵場所として認識しているとしています(写真12参照).さらにこの論文の中で彼は生息地選択に関する過去の研究をレビューして,「トンボの生息地選択は基本的に視覚によって行われることが仮定されており,物理・化学的刺激が視覚判断をより確固としたものにするだろう.そして産卵場所の選択には,視覚と触覚が非常に重要である」と述べています.
視覚が生息地選択で重要な役割をしていることを直接に証明することは困難なことと思われますが,視覚の重要性を示す実験的研究があり,否定的な見解はほとんど見られないことから,生息地選択に視覚を利用していることは,ほとんど間違いのないことであると思われます.もちろん視覚以外の感覚器官を使っている可能性を否定することはできませんが.
*3.選択実験
choice experiments
写真12.:水面に太陽光が反射する方向で撮影した池(左)と水田(右).いずれも黒っぽい背景に反射光が輝いている状態になる.
複眼の各個眼には虹彩色素細胞があり入ってくる光の量を調節できるので,まぶしい光が入ると背景は写真のように黒っぽくなると思われる.
トンボの生態学 2.生息地選択
トンボの成虫群集と関連ある環境要素
ある生息場所のトンボ群集は,それらのトンボが生息地選択を行った結果できた集団です.したがって,トンボ群集を解析すれば,生息地選択に関する情報が得られる可能性があります.
トンボは種によって異なる環境要求性があります
*1.しかし異なるといっても,その相違の程度はさまざまで,かなり似ているものから相当に違うものまでいろいろとあるはずです.当然の結果として,環境要求性が似ているものは似たような生息場所を選ぶことになり,相違の程度が大きいものは異なる生息場所を選ぶことになるでしょう.その結果,環境の異なるさまざまの生息場所には,それぞれ独特のトンボ群集が形成されることになります.
この考え方を基本原理とすれば,次のようなアイデアが浮かんできます.環境の異なる複数の生息場所(水域)を選び,それぞれの生息場所のさまざまな環境要素を数値化したデータと,それぞれの生息場所のトンボ群集(種構成や個体数)を数値化したデータとを比較することで,トンボ群集の相違に相関した環境要素の相違(環境勾配)を見いだすことができる,というアイデアです.こういったアイデアを実現する手法を多変量解析と呼びます.
私は,2001年から2003年にかけて,国立環境研究所の生物多様性プロジェクトの一環として,兵庫県南部に分布するため池のトンボ群集の調査に参加させてもらいました.その結果は,「ため池とその周辺環境における生物多様性の維持機構とその減少要因について」と題して,国立環境研究所(2006)の特別研究報告「生物多様性の減少機構の解明と保全プロジェクト(終了報告)平成13~17年度」の中に掲載されています
*2.ここではこの結果を,トンボの生息地選択という視点で見直して,お話を進めていきましょう.
兵庫県南部播磨地域に分布する,環境の異なる35のため池を選び,それらため池の物理化学的および生物的環境要素,またため池周辺の土地利用の状態を数値化しました.トンボ群集(種と個体数)は,生物季節的に全ての種を網羅できるよう,成虫年6回(春,晩春,初夏,盛夏,初秋,秋),幼虫年3回(青木,2004参照)の調査を,それぞれのため池で行いました.そしてその結果を多変量解析の一つ,DCAにかけて分析しました.その結果が図2です.
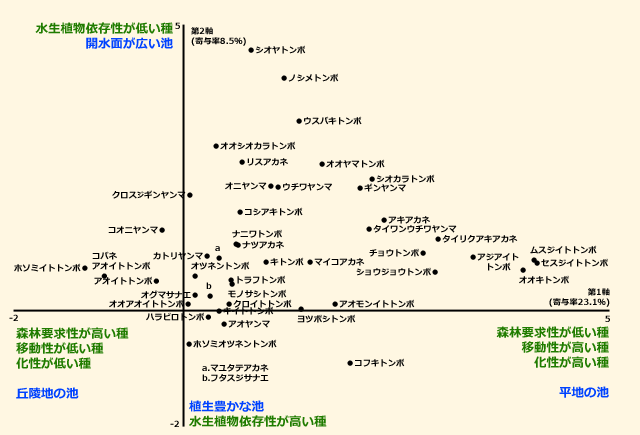
図2.兵庫県南部地方の35のため池のトンボ群集のDCA (国立環境研究所,2006をもとに作成)
第1軸右側には平地の皿池に生息するような種が,左側には谷池に生息するような種が来た.
第2軸の上側には産卵基質として水生植物が不要な種が,下側にはそれが必要な種が来た.
青字:環境勾配,緑字:トンボの生態的勾配.
図2は,トンボ各種が,ため池の有する何らかの環境勾配に沿って配置されていると考えることができます.その環境勾配は,DCAから得られた各ため池のプロットを,図2に重ね合わせて表示することで明らかになります(プロットは省略して環境勾配だけを図2に表示).
数値化された数多くの環境要素のうち,第1軸に沿ったため池の並びと有意に相関のあったものを手元の資料
*3でまとめると,正(右)の方向には,池の周囲長が大きく,隣接地の起伏の程度が低く,200m内の森林面積が小さく,200m内の水田面積が大きく,池周囲が開けていて日射量が多いなど,平地的環境の中にあるため池(いわゆる皿池)が位置します.逆に負(左)の方向には,池のサイズが小さく,隣接地の起伏の程度が高く,200m内の森林面積が大きく,200m内の水田面積が小さく,池の日射量が少なく,特定の方角に陰ができる傾向が強い鬱閉的な池であるなど,丘陵地的環境の中にあるため池(いわゆる谷池)が位置します.
一方,トンボ群集の生態的特性は,図2にあるように,負(左)の方向にはまず森林要求性の高い種が来ています.環境要素としては200m内の森林面積が大きいため池が来ていますので,生活史のある時期に隣接する森林を利用しているトンボたちが多く来ていることが分かります.例えばアオイトトンボは未熟な時期に近くの林床にもぐり込みます.移動性に関しては,隣接した森林を利用して生活しているため,それらのトンボたちにはあまり必要がありません.化性が低いということは,幼虫期間が長いということで,羽化までに2,3年かかる種が含まれているということです.長期間幼虫で過ごすためには水が枯れることがあってはなりませんが,森林は水を供給しますので,その可能性は低くなります.
正(右)の方向はその逆で,セスジイトトンボのような周辺の草地と池で一生を送るような,生活史の中に特に森林を必要としない種が来ています.したがって,これらは隣接森林の少ない平地の池で生活できます.またオオキトンボやアキアカネのように移動性がある種は,もし森林で生活する期間があったとしても,それが隣接して存在する必要はなくなります.平地の池で生活するトンボたちは幼虫期間が短いものが多いのですが,これは,これらがかつて氾濫原に生活していたトンボたちだったと考えられ,一般に水が枯渇しやすい環境に対する適応であると考えられます.その結果化性が高くなっているのでしょう.
周辺環境要素の中で,トンボ群集と関わりの深いものとして,「森林面積や水田面積の範囲が200m内」という数値が出ています.これについては,Corbet (1980) にもその数値が出ていて,いくつかの研究を引用して次のように述べています.「成虫が午後から夕方に戻っていくねぐらは,雌雄の出会い場所
*4から1~200m程度の範囲にある密に茂った叢林や草地である」.つまり,この調査結果の「200m内の範囲にある樹林や水田」というのは,日周活動においてねぐらとしても利用されているということです.なお「出会い場所」については,本調査で出現した種のほとんどは,水域(ため池)が出会い場所になっています.
*1.種によって異なる生態学的ニッチをしめているともいえるだろう.
*2.この冊子はインターネットで
pdfファイルが手に入るので,方法や結果の詳細はそれを読んでください.
*3.国立環境研究所(2006)では,互いに相関する環境要素は省略されていて,DCA各軸と最も高い相関関係のあるものだけを表示しています.このDCA結果のデータは筆者も有しており,本文では,生息地選択の文脈で必要と思われる環境要素を,互いに相関していても抽出しています.例えば,池の面積が大きいほど池の日射量は大きくなるので,池の面積と日射量は互いに高い相関関係があります.ここではこの両者をともに取り上げています.それはトンボの生息地選択に関わる環境のリリーサーが,池の大きさか日射量かは分からないからです.
*4.出会い場所 rendezvous
写真13.DCA第1軸の環境勾配のイメージ:(左)第1軸左側の,森林が隣接した丘陵地の池.(右)第1軸右側の,水田や草地に囲まれた平地の池.
同様に,第2軸に沿ったため池の並びと有意な相関のあった環境要素は,水生植物の種数・多様性・被度などが負(下)の方向に,その反対の開水面の広さ・割合が正(上)の方向に出ました.トンボ群集の生態的特性としては,産卵管を持ち,植物組織内に産卵し,幼虫が植生内生活をするものが負(下)の方向に多く現れ,生殖弁を持ち,植物外の基質(水面,泥面,浮遊物,空中)に産卵し,幼虫が底生生活をするものが正(上)の方向に多く現れました.これらは産卵に関わる環境要素であるといえます.
以上のように,トンボ群集の生態的勾配とため池の環境勾配の間には,意味ある相関関係が成り立っていることが分かります.すなわち,寄与率の最も高い第1軸はトンボ成虫の生活史や日周活動のような生存に深く関わる環境要求を満たす環境勾配であり,次に高い第2軸はそのうち水域で行われる産卵のような繁殖に関わる環境要求を満たす環境勾配であるとまとめられます.
写真14.DCA第2軸の環境勾配のイメージ:(左)第2軸上側の,開水面が多く植生が少ない池.(右)第2軸下側の,植生豊かな池.
この調査研究以外にも,成虫群集と環境要因の関係について,海外で多変量解析等の手法を使って研究がなされてきています.多くの研究で共通して抽出された要因は,日射(日陰)の多少 (例えば Steytler & Samways, 1995; Samways et al., 1996; Clark & Samways, 1996; Osborn & Samways, 1996; Stewart & Samways, 1998; Kinvig & Samways, 2000; Oertli et al., 2002),および水生植物の多少 (例えば Buchwald, 1992; Steytler & Samways, 1995; Samways et al., 1996; Clark & Samways, 1996; Osborn & Samways, 1996; Stewart & Samways, 1998) の2つで,これらの環境傾度が成虫群集の違いを大きくしているという結果が示されています.その他には,池のサイズの大小 (Oertli et. al, 2002),川の流速 (Steytler & Samways, 1995),周辺の草本及び樹林の有無や多少 (Steytler & Samways, 1995; Stewart & Samways, 1998),水域の恒久性あるいは一時性 (Clark & Samways, 1996) などの要因によって,トンボ群集が大きく違ってくると報告されています.これらの環境要素は,このため池調査結果で得られたものと多くが一致しています.
トンボの生態学 2.生息地選択
フィルターモデル
上記で検討したトンボ群集と深く関係する環境要素は,すべて視覚で判断できるものです.というより,隣接森林の有無,池のサイズ,日当たりの具合,水生植物の多少など,視覚を使わずしてどうして判断できるのでしょう.視覚以外の感覚器官によって得られた情報によってトンボ群集が形成され,これらの環境要素がそのその情報と高い相関があった結果,偶然にこのような結果になったという論理も可能かもしれませんが,現時点ではまったくその証拠はありません.こういったことから,以下では,トンボがこれらの環境要素に視覚で反応して群集を成立させていると仮定して,話を進めていきたいと思います.
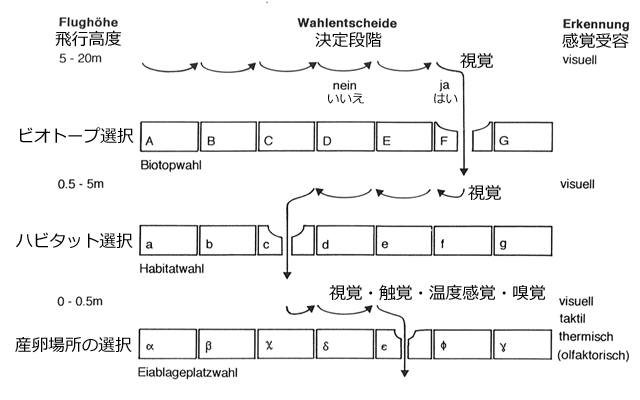
ため池のDCA結果から,寄与率の高い第1軸の環境勾配は,池全体のサイズとか,200m内の森林や水田とか,あるいは池の日当たりがよいか悪いかなど,ため池やその周辺というかなり大きな環境要素(ビオトープレベル)に反応していることが分かります.それに対して寄与率が次に高い第2軸の環境勾配は,水生植物の被度や多様性という,より小さな環境要素(ハビタットレベル)に反応していることがわかります.Samways et al.(1996) も,種と環境の関係は,止水性種でも,流水性種でも,微小生息場所よりは空間的スケールの大きなビオトープレベルにより強い結びつきがある,と述べています.これらの関係性は,フィルターモデル (Wildermuth, 1994) の考え方としてまとめられます.
図3.フィルターモデル.Wildermuth (1994) に訳語を加筆.
トンボが最終的に産卵場所を選択・決定するまでの判断手順.
A~G,a~g,α~γ はそれぞれ質の異なる,ビオトープ,ハビタット,産卵場所を示す.
フィルターモデルを使うと,トンボが産卵場所にやって来る手順は,やや擬人的になりますが,次のように説明することができます.まず5~20mくらいの高さを飛び,視覚を使って反射光などの情報からそこに水があることを確認します.さらに周辺の景観(森林があるか草地か),池の日当たり状況などを判断します.自分に適していない場合は飛び去って別の場所を探索します.そして自分に適している場所を見つけると(図3のF),高度を下げてより小さな環境要素を視覚で探索します.水生植生の構造や状態,水際や岸辺の構造など,産卵に適した状態や構造を持っているかどうかを,その水域のあちこちを飛び回って検査します.それがなければ飛び去って別のビオトープを探しに行きます.もしその水域内に適した産卵場所の構造を見つければ(図3のc),さらに降下したり産卵基質の上に止まったりして,それを検査します.最終的に産卵に適しているかどうかは,触覚(産卵基質に触れる動作)や,水温(打水動作)などの感覚によって決定されます.そして適していると判断したら(図3のε),そこで産卵を始めます.
以上はメスの話ですが,水域で繁殖活動をするトンボのオスも,同様の手順でハビタットまたは産卵場所の選択を行っていると思われます.観察に出かけると,不思議なくらいオスはメスが来る場所を知っています.これは少なくともハビタットの選択レベルまではメス同様に行われているからと考えられます.さらに,オスがときどき打水動作をすることがありますが,これはひょっとすると,産卵基質の検査をしている行動かもしれません.
フィルターモデルの典型的な行動は,アキアカネの産卵活動で観察することができます.秋の朝,アキアカネはタンデムになって10~20mの高さを飛び,ねぐらから産卵場所へ移動します.そして,アキアカネの産卵に適した水田,すなわち水がたまって湿地状になった稲刈り跡の水田群を見つけると,高度を下げてもっともよい状態のところを探します.そして,ここと決めたところで産卵を開始します.この最後の2段階では,少し産卵しては飛び去るなどの,試行錯誤をしている状況も観察できます.

写真15.フィルターモデル,ビオトープの選択:アキアカネが上空20mあたりを飛んで(左),水田地帯を探索している(右).
写真16.フィルターモデル,ハビタットの選択:アキアカネが地上3mあたりを飛んで(左),水のたまった水田を探している(右).
写真17.フィルターモデル,産卵場所の選択:アキアカネが産卵場所の直上を飛んで(左),実際に産卵活動を行っている(右).
以上は,環境の刺激に対して,トンボが主に視覚で反応するという文脈で説明してきましたが,水が伏流していて見えないところをオスがパトロールしたり,水がなくなった湿地で産卵するトンボがいたりと,水を視覚的にとらえることができない場所で繁殖行動をするトンボの存在があって,別の感覚器官の情報が生息地選択に使われている可能性は捨てきれません.またトンボが集団で産卵している状況が誘引効果になる可能性も指摘されていて,これは環境要素に対する視覚的反応より強いかもしれません.
ただ,水のないところで産卵する形質については,ノシメトンボで面白い報告があります.ノシメトンボは,通常,水を落とした水田や水が引いた草地で産卵します(写真18左参照).Watanabe & Kato (2012)は,稲穂の上で産卵するノシメトンボについて,高さのそろった稲の上面から水蒸気が空気中に立ち上り,その分圧が複数の観察日でよく似た値を示しており,かつ稲上面から高くなるほど低くなっていくことを示した上で,その水蒸気によって生じる水平偏光が,ノシメトンボの産卵を誘引するということを述べています.
私のフィールドで観察しているネアカヨシヤンマは,完全に水の涸れた天水湿地で,よく晴れた日には決まって午後2時に産卵にやってきます(写真18右参照).これを複数回確認しています.これも湿土から立ち上る水蒸気によって生じる偏光が起因しているのかもしれません.夏の午後2時というのはもっとも気温が高くなる時間帯で,そのときには激しく水蒸気が立ち上り,地面付近の水蒸気分圧も高くなり,水平偏光の割合が高くなっている可能性があります.
最後に,ここではまったく議論しませんでしたが,トンボの行動が単純な刺激-反応系で成り立っているとすれば,ここで議論したさまざまの環境要素中のいったい何が,生息地に集まるという反応を引き起こす刺激(cue)になっているのかということについては,ほとんど分かっていません*1.まだまだ生息地選択に関しては研究していく分野が残されています.
*1.水面からの反射光は,反応を引き出す刺激になっている可能性が指摘されている(Wildermuth & Spinner, 1991).
写真18.水のないところで産卵するトンボ.(左)ノシメトンボ:水の引いた草地で産卵.(右)ネアカヨシヤンマ:水が涸れた天水湿地で産卵.
参考文献
青木典司,1997.標識調査によるベッコウトンボ成虫の動態について.昆虫と自然 32(7):11-18.
青木典司,2004.トンボ類の多様性調査法.昆虫と自然 39(14):13-14.
青木典司,角野康郎,三橋弘宗,田中哲夫,村上俊明,中川恵,高村典子.2002.ため池から生物多様性の保全を考える (4)トンボ群集を決める要因.日本陸水学会第67回大会講演要旨集:193.
*新井 裕,1989.オジロサナエの幼虫は川を下るか.インセクタリウム 27:84-87.
新井 裕,1993.オジロサナエの生活史.Tombo 36(1/4):29-34.
Buchwald, R., 1992. Vegetation and dragonfly fauna - charasteristics and examples of biocenological field studies. Vegetatio. 101(2): 99-107.
Clark T.E. & Samways M.J., 1996. Dragonflies (Odonata) as indicators of biotope quality in the Kruger National Park, South Africa. J. Appl. Ecol. 33(5):1001-1012.
Corbet P.S., 1980. Biology of Odonata. Ann. Rev. Entomol. 25:189-217.
Corbet P.S., 1999. Dragonflies Behavior and Ecology of Odonata. Cornell Universitiy Press. New York.
浜田康・井上清,1985.日本産トンボ大図鑑.講談社,東京.
可児藤吉,1944.渓流性昆虫の生態.古川晴男編「昆虫」上,研究社,東京.
Kinvig R.G. & Samways M.J., 2000. Conserving dragonflies (Odonata) along streams running through commercial forestry. Odonatologica 29(3): 195-208.
国立環境研究所,2006.生物多様性の減少機構の解明と保全プロジェクト(終了報告) 平成13~17年度.国立環境研究所特別研究報告, SR-72-2006, 81pp.
連 日清・松木和夫,1979.台湾産ハラビロトンボ2種の幼虫について.昆虫と自然 14(6):57-60.
三時輝久・平田真二,1997.ベッコウトンボの移動習性.昆虫と自然 32(7):27-32.
水野信彦・御勢久右衛門,1993.河川の生態学 補訂新装版,沼田真編.築地書館,東京.
Obson R. & Samways M.J., 1996. Determinants of adult dragonfly assemblage patterns at new ponds in south Africa. Odonatologica 25(1): 49-58.
Oertli B., Joye D.A., Castella E., Juge R., Cambin D. and Lachavanne J.B., 2002. Does size matter? The relationship between pond area and biodiversity. Biol. Conserv. 104(1): 59-70.
Samways M.J., Caldwall P.M. & Obson R., 1996. Spatial patterns of dragonflies (Odonata) as indicators for design of a conservation pond. Odonatologica 25(2): 157-166.
*Schwind, R., 1991. Polarization vision in water insect and insects living in moist substrate. J. Comp. Physiol. A169:531-540.
Stewart D.A.B. & Samways M.J., 1998. Conserving dragonfly (Odonata) assemblages relative to river dynamics in an African savanna game reserve. Conserv. Biol. 12(3): 683-692.
Steytler N.S., & Samways M.J., 1995. Biotope selection by adult male dragonflies (Odonata) at an artificial lake created for insect conservation in South Africa. Biol. Conserv. 72(3): 381-386.
杉村光俊・石田昇三・小島圭三・石田勝義・青木典司,1999.原色日本トンボ幼虫成虫大図鑑.北海道大学図書刊行会.札幌.
Watanabe, M. & K. Kato, 2012. Oviposition behaviour in the dragonfly Sympetrum infuscatum (Selys) mistaking dried-up rice paddy fields as suitable oviposition sites (Anisoptera: Libellilidae). Odonatologica 41(2):151-170.
*Wright, M. and Shoup, C.S., 1945. Dragonfly nymphus from the Obey River drainage and adjacent streams in Tennessee. J. Tenn. Acad. Sci. 20:226-278.
Wildermuth, H., 1994. Habitatselektion bei Lebellen. Advances in Odonatology. 6:223-257.
Wildermuth, H., & Spinner, W., 1991. Visual cues in oviposition site selection by Somatochlora arctica (Zetterstedt) (Anisoptera: Corduliidae). Odonatologica 20:357-367.
山本哲央・新村捷介・宮崎俊行・西浦信明,2009.近畿のトンボ図鑑.いかだ社,東京.