最近新聞などで,南方のチョウが近畿地方や北陸地方で毎年のように見つかり温暖化によるものだなどという記事がよく載ります.イシガケチョウやナガサキアゲハなどかなりの種類にのぼるのではないでしょうか.トンボでもこれと同じように,南方種が北上しているのではないかとかなり以前から言われています.その中で最近目立って分布を北の方へ拡大しているのが,タイワンウチワヤンマとベニトンボです.ここではこのうち,タイワンウチワヤンマの分布北上についてお話ししたいと思います.
神戸のトンボ/トンボの話題-温暖化? 北上するタイワンウチワヤンマ 改訂版
更新:2022.02.06 12:00
更新:2022.02.06 12:00
温暖化? 北上するタイワンウチワヤンマ 改訂版
北上する昆虫たち

写真1.1990年から神戸でみつかり始めたタイワンウチワヤンマ.1993.8.29.,兵庫県神戸市.

写真2.左:最近北上傾向が著しいベニトンボのオス,2013.7.7., 徳島県美波町.
右:温暖化で北上しているといわれるナガサキアゲハ,2007.5.4., 兵庫県小野市.
右:温暖化で北上しているといわれるナガサキアゲハ,2007.5.4., 兵庫県小野市.
片谷(2017)によると,タイワンウチワヤンマは,ネパール,インド,ミヤンマー,ラオス,ベトナムなどの地域から,中国東部,台湾を経て日本にいたる地域に分布しており,我が国がその北限となっています.ですから,日本でのタイワンウチワヤンマの北上は,そのまま本種の北限記録の更新となります.
温暖化? 北上するタイワンウチワヤンマ 改訂版
記録から見たタイワンウチワヤンマの分布拡大の様相
タイワンウチワヤンマが現在の日本領土内で初めて記録されたのは19世紀で,場所は『琉球(Loo-Choo)』となっています(Selys, 1888).詳細なロケーションは不明ですが,この時期にすでに南西諸島にいたことがわかります.そして,いわゆる本土(北海道・本州・四国・九州)で記録されたのは1934年の高知県安芸郡田野町(佐々木,1936)が最初で,1938年には鹿児島県祁答院町と宮崎県宮崎市(朝比奈,1957)でも記録されています.当時はトンボの調査をする人はとても少なかったと思われますから,この記録から四国の方に先にやってきたとはいえませんし,またそれ以前から本土にいたかどうかもわかりません.しかし本種の分布拡大を考える際の,貴重な出発点となる記録であることは間違いありません.
そしてその後の記録を追いかけていくと,1971年の三重県北牟婁郡紀北町(旧紀伊長島町)(冨田,1971)の記録を除けば,ほとんどすべての初発見記録が,これらの出発点から時間的・地理的に順序よく並びます.そこで,まず各地域の初記録を追いかけて,分布拡大の経路を推定してみたいと思います.なお市町村名は文献に掲載されたものを現在の新地名に変更しています.
そしてその後の記録を追いかけていくと,1971年の三重県北牟婁郡紀北町(旧紀伊長島町)(冨田,1971)の記録を除けば,ほとんどすべての初発見記録が,これらの出発点から時間的・地理的に順序よく並びます.そこで,まず各地域の初記録を追いかけて,分布拡大の経路を推定してみたいと思います.なお市町村名は文献に掲載されたものを現在の新地名に変更しています.
各地域の詳細な分布拡大の様相について,特に見る必要のない方はまとめへジャンプしてください.
→まとめへジャンプ
→まとめへジャンプ
Ⅰ.九州の分布拡大
九州においては,1938年の祁答院町と宮崎市をスタートとして,まず西岸沿いに,1957年に長崎県雲仙(池崎,1961),1959年に熊本県金峯山(舛田,1959),1965年に福岡県久留米市(池田,1969),1967年に佐賀県佐賀市富士町(佐賀北高校生物部昆虫班,1967),1971年に福岡県福岡市・長崎県佐世保市(私信),1972年に佐賀県唐津市(市場,1973b)・長崎県松浦市(池崎,1977),1993年に福岡県北九州市八幡西区(久川,1994)とほぼ九州の北端に達し,1995年には海を渡って長崎県壱岐(井上・相浦,1995)でみつかり,2007年には対馬に生息していることが新聞に載りました(長崎新聞,2010年8月13日).
一方東岸沿いの記録は非常に少なく,1938年の宮崎市の記録の後は,1970年に大分県宇佐市(佐藤,1980)の記録が出るまで,この間の記録が見当たりません.ただ,1972年に延岡市にいることが報告されており(永井,1974),東岸沿いに分布は広がっているようです.その後は1972年に福岡県田川市(私信),1978年に大分県中津市(平松,1979),1992年に福岡県苅田町(松木,1993)と初記録が北進しています.
九州においては,1938年の祁答院町と宮崎市をスタートとして,まず西岸沿いに,1957年に長崎県雲仙(池崎,1961),1959年に熊本県金峯山(舛田,1959),1965年に福岡県久留米市(池田,1969),1967年に佐賀県佐賀市富士町(佐賀北高校生物部昆虫班,1967),1971年に福岡県福岡市・長崎県佐世保市(私信),1972年に佐賀県唐津市(市場,1973b)・長崎県松浦市(池崎,1977),1993年に福岡県北九州市八幡西区(久川,1994)とほぼ九州の北端に達し,1995年には海を渡って長崎県壱岐(井上・相浦,1995)でみつかり,2007年には対馬に生息していることが新聞に載りました(長崎新聞,2010年8月13日).
一方東岸沿いの記録は非常に少なく,1938年の宮崎市の記録の後は,1970年に大分県宇佐市(佐藤,1980)の記録が出るまで,この間の記録が見当たりません.ただ,1972年に延岡市にいることが報告されており(永井,1974),東岸沿いに分布は広がっているようです.その後は1972年に福岡県田川市(私信),1978年に大分県中津市(平松,1979),1992年に福岡県苅田町(松木,1993)と初記録が北進しています.
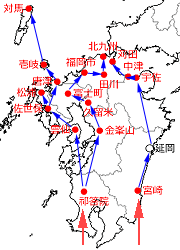
図1.九州の初記録地と近接記録地を,地理的連続性も考慮して時期順につないだもの(ただし必ずしも分布拡大経路を示しているわけではない.以下の図も同様である).
Ⅱ.四国・中国地方の分布拡大
まず四国東部では,1936年の田野町の記録に続いて,1950年代にはすでに徳島県阿南市にいたとの伝聞記述があります(平井,1980).続いて1967年に徳島県鳴門市(平井,1968),1970年に同吉野川市川島町(平井,1971),1972年に同美馬市脇町(平井,1973),1979年に同三好市池田町(平井,1980)と,吉野川を遡るように初記録が続き,1980年に池田町の北西に位置する香川県観音寺市古河町の初記録(石川,1982)へと続きます.
この池田町と観音寺市の記録の関連性に関して,石川(1983)が,観音寺市大野原町および隣接する三豊市山本町における1983年の記録を報告した際に,「(香川)県東部では(1983年現在)まだ発見できていない」と高木氏の私信を引用していること,また愛媛県立博物館研究報告第10号(楠・桑田,1979)には「1978年時点で愛媛県内にタイワンウチワヤンマの記録がない」という記述があることなどから,この観音寺市の記録につながる記録はこの池田町のものしかありません.ただ池田町と観音寺市の間には標高550mの猪ノ鼻峠という障壁が存在しています.これについては,六甲山地(神戸市)にある標高350mほどの山中の池での記録(トンボ歳時記 2009.8.24.)があって,森林とアクセス道路しかないような山中の池へも移動することが確認されていますので,この程度の峠を越えることは不可能ではないと思われます.
香川県東部の初記録は1984年のさぬき市寒川町(高木,1986)で,海を渡って1985年の池田町(小豆島)(高木,1987)へと一気に記録が北進しています.ただし鳴門市で確認されたのが1967年ですので,ここには大きなタイムラグが存在しています.これは調査の空白が原因しているかも知れません.
まず四国東部では,1936年の田野町の記録に続いて,1950年代にはすでに徳島県阿南市にいたとの伝聞記述があります(平井,1980).続いて1967年に徳島県鳴門市(平井,1968),1970年に同吉野川市川島町(平井,1971),1972年に同美馬市脇町(平井,1973),1979年に同三好市池田町(平井,1980)と,吉野川を遡るように初記録が続き,1980年に池田町の北西に位置する香川県観音寺市古河町の初記録(石川,1982)へと続きます.
この池田町と観音寺市の記録の関連性に関して,石川(1983)が,観音寺市大野原町および隣接する三豊市山本町における1983年の記録を報告した際に,「(香川)県東部では(1983年現在)まだ発見できていない」と高木氏の私信を引用していること,また愛媛県立博物館研究報告第10号(楠・桑田,1979)には「1978年時点で愛媛県内にタイワンウチワヤンマの記録がない」という記述があることなどから,この観音寺市の記録につながる記録はこの池田町のものしかありません.ただ池田町と観音寺市の間には標高550mの猪ノ鼻峠という障壁が存在しています.これについては,六甲山地(神戸市)にある標高350mほどの山中の池での記録(トンボ歳時記 2009.8.24.)があって,森林とアクセス道路しかないような山中の池へも移動することが確認されていますので,この程度の峠を越えることは不可能ではないと思われます.
香川県東部の初記録は1984年のさぬき市寒川町(高木,1986)で,海を渡って1985年の池田町(小豆島)(高木,1987)へと一気に記録が北進しています.ただし鳴門市で確認されたのが1967年ですので,ここには大きなタイムラグが存在しています.これは調査の空白が原因しているかも知れません.
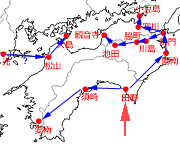
図2.四国の初記録地と近接記録地を時期順につないだもの
一方四国西部では,まず1956年に高知県須崎市(中村,1957)という記録があり,1979年愛媛県愛南町(旧御荘町)に多産するという記録へと続きます(杉村,1981).続く愛媛県の記録は1981年の松山市(窪田,1982)になりますが,旧御荘町からは山地をはさんで直線距離で100km以上離れており,海岸線伝いだとさらに距離が伸びるので,距離と時間から考えてこれら2つの記録は関連性が低いと思われます.したがって,松山市への進入は別ルートを考える必要がありそうです.その一つとして,山口県から海を渡ってきた可能性が考えられます.すなわち,1973年に記録のある山口県光市あるいは柳井市から,屋代島,情島,津和地島,怒和島,中島,睦月島,興居島と小さな島伝いに移動してきた可能性です(図3).この場合,距離は半分ほどで,時間も十分にあります.そしてその後は記録が北上し,1988年に越智郡吉海町(大島)で,すでに多数が生息していたことが確認されています(井上,1988).
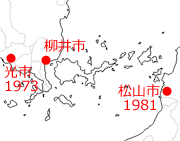
図3.光市と松山市の間の島々(CraftMAP)
http://www.craftmap.box-i.net/map.php
では続いて中国地方の記録を見ていきましょう.中国地方の初記録は山口県で,1967年の周南市徳山(池田,1967)の記録です.この時点ですでに20頭あまりの個体がいたと書かれています.これは偶然だと思われますが,この徳山と,同じく1967年に初記録が出た徳島県鳴門市は,緯度がほとんど同じになっています.ところでこの徳山への進入経路はちょっと謎に包まれています.これに関して池田(1969)は,「1968年現在福岡県久留米市が分布北限である」として,この久留米市のものが,未確認の中継地を経て山口県に入ってきたとの考えを述べています.しかし大分県宇佐市に1970年に記録があることから,それ以前のある時期に,宮崎方面から北上してきた個体が国東半島から対岸の徳山へ渡海したという可能性が考えられます.この間には姫島,野島といった小さな島があり,海を渡る距離は25km程度です.ただこの推定の根拠になる九州東岸沿いの記録が欠落しているのが残念です.
さてその後山口県東西に記録が広がります.西へは,1968年の山口市阿知須町,1969年の小野田市へと続き,その後の記録が手元にありませんが2010年には下関市にいる(私信)ことが確認されています.東へは,1969年に下松市,1973年に光市の記録があり,1969年以降の記録(時期は不記載)として柳井市,岩国市に分布することが,1988年の文献に記されています(池田,1988).
次に広島県です.初記録は瀬戸内海に浮かぶ上島(豊田郡大崎上島)での1989年で,すでに多数が生息していたと記録されています(藤原・安達,1989).藤原氏ら(1989)は,これが広島県初記録であると明確に述べていますので,広島県への進入はここから始まったと考えるべきでしょう.この島は,1988年に記録された愛媛県越智郡吉海町(大島)の北西に,大三島を挟んで隣接する島で,この2つの個体群の関連性は高いと思われます.一方この2つの個体群がどこから来たかについてははっきりとしません.近隣の記録としては,一つは愛媛県松山市(1981年),もう一つは山口県岩国市(1988年以前)のものがあります.さてその後,1992年に呉市安浦町(椎木,1993),1995年に福山市(安達ら,1996)へと東西への広がりが見られます.そして2013年に東広島市鏡山(青山ら,2014),最近の三原市久井町(柳澤,私信)などの記録があって,少し内陸部へと分布が広がっています.
さてその後山口県東西に記録が広がります.西へは,1968年の山口市阿知須町,1969年の小野田市へと続き,その後の記録が手元にありませんが2010年には下関市にいる(私信)ことが確認されています.東へは,1969年に下松市,1973年に光市の記録があり,1969年以降の記録(時期は不記載)として柳井市,岩国市に分布することが,1988年の文献に記されています(池田,1988).
次に広島県です.初記録は瀬戸内海に浮かぶ上島(豊田郡大崎上島)での1989年で,すでに多数が生息していたと記録されています(藤原・安達,1989).藤原氏ら(1989)は,これが広島県初記録であると明確に述べていますので,広島県への進入はここから始まったと考えるべきでしょう.この島は,1988年に記録された愛媛県越智郡吉海町(大島)の北西に,大三島を挟んで隣接する島で,この2つの個体群の関連性は高いと思われます.一方この2つの個体群がどこから来たかについてははっきりとしません.近隣の記録としては,一つは愛媛県松山市(1981年),もう一つは山口県岩国市(1988年以前)のものがあります.さてその後,1992年に呉市安浦町(椎木,1993),1995年に福山市(安達ら,1996)へと東西への広がりが見られます.そして2013年に東広島市鏡山(青山ら,2014),最近の三原市久井町(柳澤,私信)などの記録があって,少し内陸部へと分布が広がっています.
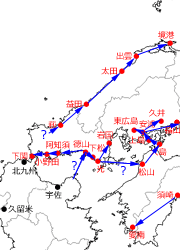
図4.四国・中国西部の初記録地と近接記録地を時期順につないだもの
岡山県の初記録は,1979年の倉敷市児島でのものです(倉敷市立自然史博物館,1994).その翌年1980年には岡山市西大寺・宝伝・牛窓町に数十匹がいるとの記録があり(山崎,1992),岡山県の瀬戸内側一帯に分布が広がっています.佐藤(1993)は,「1980年当時,タイワンウチワヤンマの県下の確認は聞かず」と述べており,これら以前に進入してきたという確実な記録はないようです(1979年の児島の記録は1994年に公開されている).ところで岡山県への進入経路ですが,広島県の初記録が1989年であること,さらに兵庫県本州側の初記録が1987年であることなどを考慮すれば,岡山県へは香川県から海を越えて進入してきたと考えるのが自然でしょう.しかし香川県の瀬戸内側の最も早い記録は観音寺市の1980年ですので,わずか1年ではありますが,倉敷市の1979年という記録への時間的連続性が途切れているように見えます.しかし記録はタイワンウチワヤンマの動きを同時的に捉えているわけではありませんので,総合的に見て,香川県から進入したというのは事実とみてよいと思われます.その後2010年には,三原市から吉備中央町を経て赤磐市に至るまでの瀬戸内側ほぼ全域および内陸部への分布拡大が起きています(青野,2011).
中国地方での分布拡大で特に注目すべきは山陰地方への進出です.山口県における日本海側での初記録は2004年の萩市です(山口県立山口博物館,2006).山口県の山陽側の初記録が出てから37年後で,1993年の北九州市の記録と比べても11年後になります.その後山陰海岸に沿って記録が次々と出ていきます.2007年に島根県益田市(宮本ら,2008),2008年に同大田市(大浜,2009),2009年に同出雲市(大浜,2012),2012年に鳥取県境港市(大浜,2013)と記録が続きます.私はまだ兵庫県の日本海側では発見していませんが,進入も時間の問題のようです.
中国地方での分布拡大で特に注目すべきは山陰地方への進出です.山口県における日本海側での初記録は2004年の萩市です(山口県立山口博物館,2006).山口県の山陽側の初記録が出てから37年後で,1993年の北九州市の記録と比べても11年後になります.その後山陰海岸に沿って記録が次々と出ていきます.2007年に島根県益田市(宮本ら,2008),2008年に同大田市(大浜,2009),2009年に同出雲市(大浜,2012),2012年に鳥取県境港市(大浜,2013)と記録が続きます.私はまだ兵庫県の日本海側では発見していませんが,進入も時間の問題のようです.

図5.四国・中国東部の初記録地と近接記録地を時期順につないだもの
Ⅲ.近畿地方の分布拡大
近畿地方でタイワンウチワヤンマが初めて見つかったのは1976年で,紀淡海峡に浮かぶ和歌山県和歌山市の友が島です(竹内,1976).続いて1979年に本土側の同市加太で記録されました(村木,1980).記録の連続性から考えると,これらは徳島県鳴門市(1967年の記録)から,鳴門海峡を渡り淡路島南部・紀淡海峡を経由してやってきたと考えられます.
その後南北および東に記録が伸びていきます.初記録だけを追いかけていくと,南へは,1980年に和歌山県湯浅町(乾風,1981),1985年に同日高郡美浜町(乾風・高松,1986),1991年には田辺市に至っています(乾風,1991).さらにこの1991年には,潮岬を越えた同東牟婁郡串本町の古座駅付近に記録があります(高松,1993).これについては,高松(1993)も田辺市以南(古座に至るまで)は池が非常に少ないと記しており,後述するように三重県側から南下して来ている可能性もあります.
東へは,1987年に和歌山県岩出市の記録(高松,1988)があって紀ノ川に沿った動きを見せ,その後さらに遡って1989年の奈良県五條市の記録(久川,1991)へと続きます.そして,1997年には同橿原市,桜井市,北葛城郡などで記録があって(笹本,1998),1998年には同奈良市・生駒市など奈良県最北部にまで途中連続的に記録が出て(尾園・桜谷,2005),2002年には同最南部の下北山村(尾園・桜谷,2005)にも記録が出るなど,奈良盆地全域とその周辺に記録が広がりました.なお,奈良県への進入は,五條市ルート以外に大阪府羽曳野市(1987年に初記録;津田・山本,1988)から奈良県香芝市へのルートや,同大阪市旭区(1989年に初記録;津田・山本,1988)から奈良県生駒市へのルートなども考えられます.隣の大阪府も含めて,このあたりの記録は,線的ではなく面的に拡大しているように見えます.
北へは,1986年に大阪市住吉区(津田・山本,1988),1989年に大阪市旭区(新村,1989)と続き,翌1990年には京都府を通り越して一気に滋賀県大津市(藤本,1991)に記録が出ます.京都府の記録は少なく,2000年に奈良市のすぐ北の京田辺市や木津川市山城(田端,2001),2012年に京都市左京区南部と2018年に京都市北区(いずれも田端,私信)の記録があるだけです.その後滋賀県では1992年に草津市まで記録が北進しています(蜻蛉研究会,1998)が,その後の北進記録はないようです.
さて,もう一つの近畿地方への進入記録は淡路島から兵庫県明石市のものです.1981年に淡路島の東浦(田中,1982)が兵庫県の初記録で,1986年には同姫路市家島町男鹿島(松田,1987)にも記録が出て,1987年に本州側の同明石市で記録が出ました(二宗,1988).明石市から東には1990年に同西宮市(井上・小川,1991)に,西には1991年に同神戸市西区(青木,1991),1992年に同明石市大久保町(青木,1992),1995年に同加古川市(東,2010),1998年に姫路市(東・青木,1999),2002年には同赤穂市(東,2010)へと記録が広がりました.兵庫県南部に関しては,1997年に当時の東端の記録である加古川市平荘町以西,相生市から姫路市の間の海岸沿いの池沼をかなり精力的に調査しましたが,タイワンウチワヤンマが見つかっていません(青木・二宗,1998).したがって,可能性として,赤穂市の記録は1980年にすでに記録のあった岡山県瀬戸内市牛窓町と関係しているかもしれません.なお,兵庫県では内陸部へも分布が広がり1999年に小野市,2004年には加西市・加東市(東,2010)にまで進入しています.
近畿地方でタイワンウチワヤンマが初めて見つかったのは1976年で,紀淡海峡に浮かぶ和歌山県和歌山市の友が島です(竹内,1976).続いて1979年に本土側の同市加太で記録されました(村木,1980).記録の連続性から考えると,これらは徳島県鳴門市(1967年の記録)から,鳴門海峡を渡り淡路島南部・紀淡海峡を経由してやってきたと考えられます.
その後南北および東に記録が伸びていきます.初記録だけを追いかけていくと,南へは,1980年に和歌山県湯浅町(乾風,1981),1985年に同日高郡美浜町(乾風・高松,1986),1991年には田辺市に至っています(乾風,1991).さらにこの1991年には,潮岬を越えた同東牟婁郡串本町の古座駅付近に記録があります(高松,1993).これについては,高松(1993)も田辺市以南(古座に至るまで)は池が非常に少ないと記しており,後述するように三重県側から南下して来ている可能性もあります.
東へは,1987年に和歌山県岩出市の記録(高松,1988)があって紀ノ川に沿った動きを見せ,その後さらに遡って1989年の奈良県五條市の記録(久川,1991)へと続きます.そして,1997年には同橿原市,桜井市,北葛城郡などで記録があって(笹本,1998),1998年には同奈良市・生駒市など奈良県最北部にまで途中連続的に記録が出て(尾園・桜谷,2005),2002年には同最南部の下北山村(尾園・桜谷,2005)にも記録が出るなど,奈良盆地全域とその周辺に記録が広がりました.なお,奈良県への進入は,五條市ルート以外に大阪府羽曳野市(1987年に初記録;津田・山本,1988)から奈良県香芝市へのルートや,同大阪市旭区(1989年に初記録;津田・山本,1988)から奈良県生駒市へのルートなども考えられます.隣の大阪府も含めて,このあたりの記録は,線的ではなく面的に拡大しているように見えます.
北へは,1986年に大阪市住吉区(津田・山本,1988),1989年に大阪市旭区(新村,1989)と続き,翌1990年には京都府を通り越して一気に滋賀県大津市(藤本,1991)に記録が出ます.京都府の記録は少なく,2000年に奈良市のすぐ北の京田辺市や木津川市山城(田端,2001),2012年に京都市左京区南部と2018年に京都市北区(いずれも田端,私信)の記録があるだけです.その後滋賀県では1992年に草津市まで記録が北進しています(蜻蛉研究会,1998)が,その後の北進記録はないようです.
さて,もう一つの近畿地方への進入記録は淡路島から兵庫県明石市のものです.1981年に淡路島の東浦(田中,1982)が兵庫県の初記録で,1986年には同姫路市家島町男鹿島(松田,1987)にも記録が出て,1987年に本州側の同明石市で記録が出ました(二宗,1988).明石市から東には1990年に同西宮市(井上・小川,1991)に,西には1991年に同神戸市西区(青木,1991),1992年に同明石市大久保町(青木,1992),1995年に同加古川市(東,2010),1998年に姫路市(東・青木,1999),2002年には同赤穂市(東,2010)へと記録が広がりました.兵庫県南部に関しては,1997年に当時の東端の記録である加古川市平荘町以西,相生市から姫路市の間の海岸沿いの池沼をかなり精力的に調査しましたが,タイワンウチワヤンマが見つかっていません(青木・二宗,1998).したがって,可能性として,赤穂市の記録は1980年にすでに記録のあった岡山県瀬戸内市牛窓町と関係しているかもしれません.なお,兵庫県では内陸部へも分布が広がり1999年に小野市,2004年には加西市・加東市(東,2010)にまで進入しています.
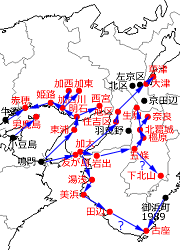
図6.近畿地方の初記録地と近接記録地を時期順につないだもの

写真3.神戸市内でタイワンウチワヤンマを初発見された岡さんが後日その発見した場所で撮影しているところ.1990.9.22.,神戸市垂水区.
Ⅳ.東海地方・関東地方の分布拡大
東海地方の最初の記録は,1971年の三重県北牟婁郡紀北町(旧紀伊長島町)のもので,発見時点ですでに幼虫も見つかっています(富田・市橋,1971).この時期タイワンウチワヤンマは徳島県鳴門市には出現していますが,まだ近畿地方には進入していません.したがってこれは海を渡って長距離飛来してきた記録と考えられます.和歌山県のトンボ研究家乾風氏は,私信で,「紀伊半島南部は台風による風力をまともに受けることから,南西方面(四国南部を指す)から蝶類の移動が知られています.トンボではウスバキトンボは早くから飛来しています.」と述べています.また吉田・八木(2012)は,「ベニトンボが(2010年に)紀伊半島南端(乾風,2011)や2011年に三重県紀宝町(毎日新聞2011年8月3日夕刊)でも採集された」ことをもとに「ベニトンボの場合もやはり四国に最も近い紀伊半島西部に飛来せず紀伊半島南端の離れた場所で発見された」と,このタイワンウチワヤンマ飛来との類似性を指摘しています.台風の通り道である紀伊半島はトンボが飛来しやすい地域であるようです.
その後紀伊半島東岸に沿って南北に記録が広がります.南へは,1985年に三重県尾鷲市(成田,1988),1989年に同南牟婁郡御浜町(下野・山口,1991),1991年に先に紹介した和歌山県同東牟婁郡串本町(古座)の記録(高松,1993)へと続き,和歌山県を南下してきた記録とつながります.一方北へは,1988年同度会郡南伊勢町(旧南島町),1991年同志摩市大王町(八木私信),そして1999年に志摩半島対岸の愛知県渥美半島先端部伊良湖国民休暇村(鵜殿,2000)の記録へと続きます.
ところですでに1988年には,遠く離れた愛知県名古屋市守山区大森湿地に記録があります(Siva-Jothy, 1997;椿私信).あまりに隔絶した記録なのと,これ以外の愛知県の記録が1999年の渥美半島先端部伊良湖国民休暇村(鵜殿,2000)が最初なので,論文の著者のSiva-Jothy博士と,当時の指導教官の椿宜高博士にメールで問い合わせました.Siva-Jothy博士は当時の状況について,「As I recall (it was a long time ago) there was always a male on the pond, and in all I must have seen 6 males that season.(思い出すと(昔のことですが)池にはいつもオスがいて、そのシーズンには全部で6匹のオスを見たに違いありません.)」と書いていて,ウチワヤンマはいたか?という私の問いに対し,明確に「No」と返答されています.また椿博士は「マイク(Siva-Jothyのこと)はもともとウチワヤンマの腹部の大きなうちわを見たかったけれど,採集したものは小さくタイワンウチワヤンマだと言われ残念がったことを覚えている.」と書いており,採集品の同定も3名の日本の研究者があたったということです.ここまで明確なお返事をいただいたこともあり,私はこの記録を確かなものとして扱っています.
さて,この名古屋守山区の記録を除いたその後の北上記録については,吉田ら(2012,2013,2014,2015,2016,2017)に非常に詳細にまとめられています.それによると,1991年の志摩市大王町を起点にして,濃尾平野を北進していき,2011年の岐阜県海津市,2016年の同羽島郡笠松町へと至っています.
東海地方の最初の記録は,1971年の三重県北牟婁郡紀北町(旧紀伊長島町)のもので,発見時点ですでに幼虫も見つかっています(富田・市橋,1971).この時期タイワンウチワヤンマは徳島県鳴門市には出現していますが,まだ近畿地方には進入していません.したがってこれは海を渡って長距離飛来してきた記録と考えられます.和歌山県のトンボ研究家乾風氏は,私信で,「紀伊半島南部は台風による風力をまともに受けることから,南西方面(四国南部を指す)から蝶類の移動が知られています.トンボではウスバキトンボは早くから飛来しています.」と述べています.また吉田・八木(2012)は,「ベニトンボが(2010年に)紀伊半島南端(乾風,2011)や2011年に三重県紀宝町(毎日新聞2011年8月3日夕刊)でも採集された」ことをもとに「ベニトンボの場合もやはり四国に最も近い紀伊半島西部に飛来せず紀伊半島南端の離れた場所で発見された」と,このタイワンウチワヤンマ飛来との類似性を指摘しています.台風の通り道である紀伊半島はトンボが飛来しやすい地域であるようです.
その後紀伊半島東岸に沿って南北に記録が広がります.南へは,1985年に三重県尾鷲市(成田,1988),1989年に同南牟婁郡御浜町(下野・山口,1991),1991年に先に紹介した和歌山県同東牟婁郡串本町(古座)の記録(高松,1993)へと続き,和歌山県を南下してきた記録とつながります.一方北へは,1988年同度会郡南伊勢町(旧南島町),1991年同志摩市大王町(八木私信),そして1999年に志摩半島対岸の愛知県渥美半島先端部伊良湖国民休暇村(鵜殿,2000)の記録へと続きます.
ところですでに1988年には,遠く離れた愛知県名古屋市守山区大森湿地に記録があります(Siva-Jothy, 1997;椿私信).あまりに隔絶した記録なのと,これ以外の愛知県の記録が1999年の渥美半島先端部伊良湖国民休暇村(鵜殿,2000)が最初なので,論文の著者のSiva-Jothy博士と,当時の指導教官の椿宜高博士にメールで問い合わせました.Siva-Jothy博士は当時の状況について,「As I recall (it was a long time ago) there was always a male on the pond, and in all I must have seen 6 males that season.(思い出すと(昔のことですが)池にはいつもオスがいて、そのシーズンには全部で6匹のオスを見たに違いありません.)」と書いていて,ウチワヤンマはいたか?という私の問いに対し,明確に「No」と返答されています.また椿博士は「マイク(Siva-Jothyのこと)はもともとウチワヤンマの腹部の大きなうちわを見たかったけれど,採集したものは小さくタイワンウチワヤンマだと言われ残念がったことを覚えている.」と書いており,採集品の同定も3名の日本の研究者があたったということです.ここまで明確なお返事をいただいたこともあり,私はこの記録を確かなものとして扱っています.
さて,この名古屋守山区の記録を除いたその後の北上記録については,吉田ら(2012,2013,2014,2015,2016,2017)に非常に詳細にまとめられています.それによると,1991年の志摩市大王町を起点にして,濃尾平野を北進していき,2011年の岐阜県海津市,2016年の同羽島郡笠松町へと至っています.
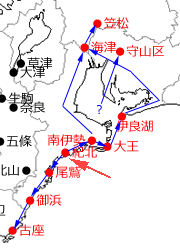
図7.東海地方西部の初記録地と近接記録地を時期順につないだもの
一方東方の初記録は,1996年に,なんと当時知られていた東限記録(Siva-Jothyの論文は1997年に公表)である三重県志摩市から100km以上離れ,大井川をかなり遡った山中にある静岡県島田市川根町で,羽化殻が70個以上も発見された(2回の調査)というものです(中岡,1996).そこで福井・加藤(1997)は,同1996年,三重県志摩市の記録との関連を調査する目的で,大井川から西へ志摩半島対岸の愛知県渥美半島にかけてタイワンウチワヤンマの分布調査を行いました.その結果,静岡県牧之原市,同御前崎市,浜松市南区倉松町でタイワンウチワヤンマの存在が確認されましたが,それより西及び渥美半島では確認ができなかったということです.その結果「(志摩半島と川根町の)中間地点である遠州灘海岸(浜松市)で記録されたことから,...(川根町の個体群は)人為的に持ち込まれたものではなく,南方から徐々に分布圏を広げてきと考えるのが妥当であると思われる」と考察しています.ただ彼らも仮定として述べていますが,遠州浜は,オオギンヤンマなど南方からの飛来種がよく見つかる場所で,タイワンウチワヤンマが遠方から海を越えて静岡県に飛来した可能性は皆無ではないでしょう※1.
その後静岡県の記録はさらに東へ延びましたが,それらより先に,2002年,神奈川県横浜市鶴見区で関東地方の初記録が出ました(石川,2002).その後2011年に同足柄上郡大井町(苅部,2013)や同小田原市(岸,2012)で記録され,2015年には東京都千代田区に初記録が出ました(石川・伊藤,2016).その後2019年に千葉県習志野市で初記録(互井・芝原,2019)が出て,これが2022年1月現在の東限となっています.面白いことに東京都の記録地の一つである立川市(互井・芝原,2019)と鳥取県の記録地境港市は,ほぼ同じ緯度(北限)になっています.
その後静岡県の記録はさらに東へ延びましたが,それらより先に,2002年,神奈川県横浜市鶴見区で関東地方の初記録が出ました(石川,2002).その後2011年に同足柄上郡大井町(苅部,2013)や同小田原市(岸,2012)で記録され,2015年には東京都千代田区に初記録が出ました(石川・伊藤,2016).その後2019年に千葉県習志野市で初記録(互井・芝原,2019)が出て,これが2022年1月現在の東限となっています.面白いことに東京都の記録地の一つである立川市(互井・芝原,2019)と鳥取県の記録地境港市は,ほぼ同じ緯度(北限)になっています.
※1.名古屋市守山区と川根村は,浜松経由で海岸を通り大井川を遡ったとすると200km以上になる(Googleのルート検索).この距離を8年(守山区が1988年,川根町が1996年にすでに多数いたので進入はそれ以前)で移動する場合,平均速度は少なくとも25km/年(また志摩市と川根村について同様に計算すると40km/年以上)となり,後に考察するように,近畿地方における陸上平均速度6~9km/年よりはるかに大きい.したがって,浜松あたりの遠州浜に,海を越えて飛来した可能性は十分にある.
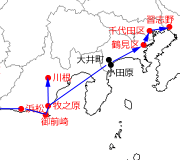
図8.東海地方東部・関東地方の初記録地と近接記録地を時期順につないだもの
Ⅴ.まとめ
以上かなり長くなりましたが,詳細に各地域の分布拡大記録を検討してきました.ご覧いただいたように,初記録地が,かなりきれいに年代を追って並んでいることにお気づきでしょう.タイワンウチワヤンマは非常に目立つトンボであり,分布拡大種として注目されてきた種ですので,初記録はニュースバリューが高く,各地の同好会誌等にほぼもれなく公表されているからだと思われます.一方,当たり前のことですが,初記録をつないだもの(図1~図8)が必ずしも分布拡大の経路を示しているわけではないことも注意しておく必要があります.
しかしそれでもなお,この見事なほどの記録の連続性は,10年,20年単位で見ていけば,発見時期の「誤差」を吸収してしまうほどの精密さを持ったものであるともいえるでしょう.そこで,20年単位で分布拡大前線を引いてみると,全国的な分布拡大状況をかなり正確にとらえることができるはずです.それが図9です.これを見ると,海岸や河川(例えば徳島県のの吉野川,和歌山県の紀ノ川など)に沿って,あるいは大きな平野では面的に,主に標高の低いところを選んで広がっていることも分かります.
以上かなり長くなりましたが,詳細に各地域の分布拡大記録を検討してきました.ご覧いただいたように,初記録地が,かなりきれいに年代を追って並んでいることにお気づきでしょう.タイワンウチワヤンマは非常に目立つトンボであり,分布拡大種として注目されてきた種ですので,初記録はニュースバリューが高く,各地の同好会誌等にほぼもれなく公表されているからだと思われます.一方,当たり前のことですが,初記録をつないだもの(図1~図8)が必ずしも分布拡大の経路を示しているわけではないことも注意しておく必要があります.
しかしそれでもなお,この見事なほどの記録の連続性は,10年,20年単位で見ていけば,発見時期の「誤差」を吸収してしまうほどの精密さを持ったものであるともいえるでしょう.そこで,20年単位で分布拡大前線を引いてみると,全国的な分布拡大状況をかなり正確にとらえることができるはずです.それが図9です.これを見ると,海岸や河川(例えば徳島県のの吉野川,和歌山県の紀ノ川など)に沿って,あるいは大きな平野では面的に,主に標高の低いところを選んで広がっていることも分かります.
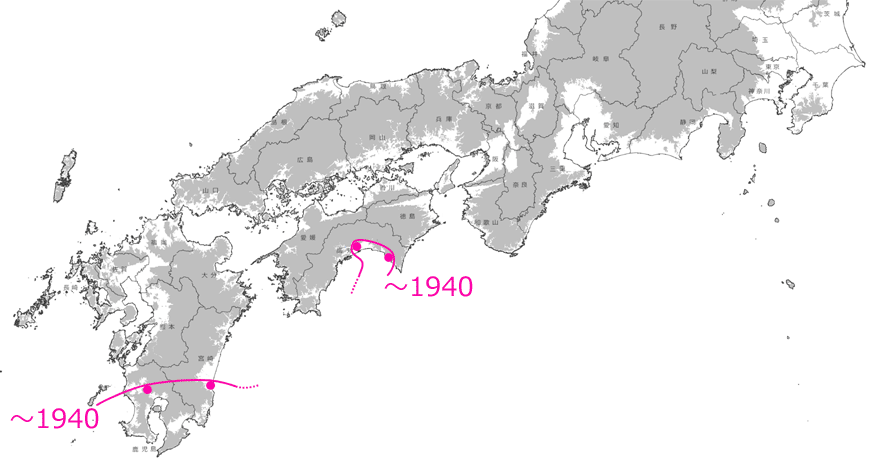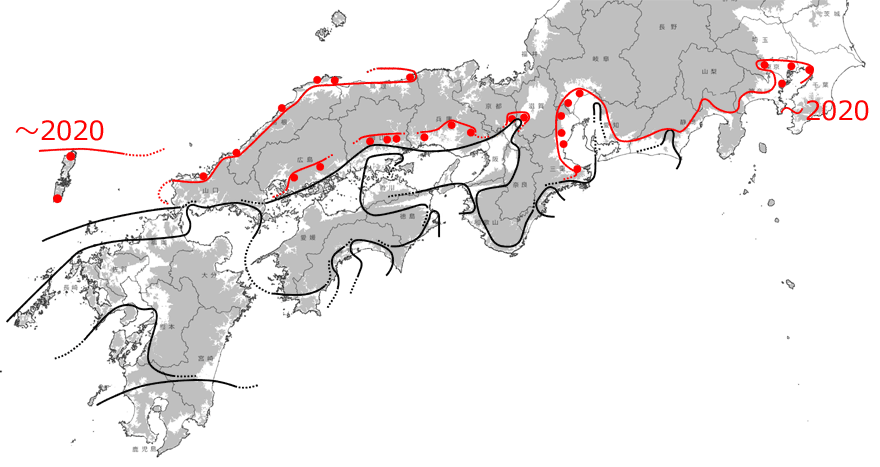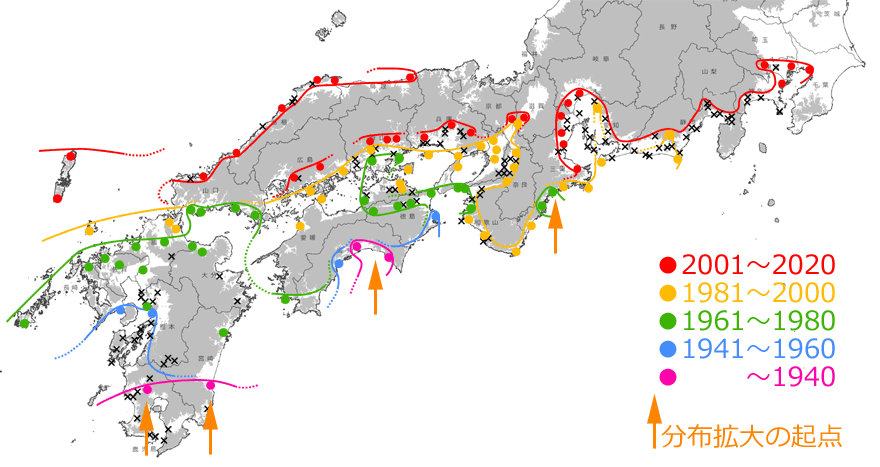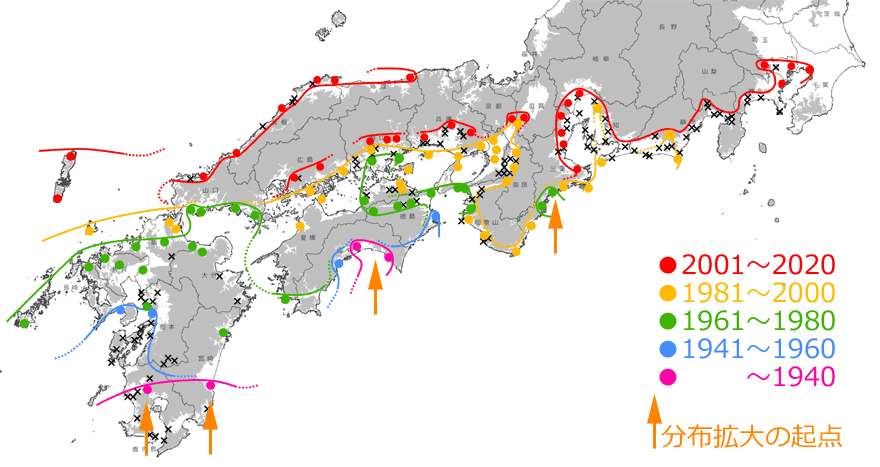
図9.20年単位で見たタイワンウチワヤンマの分布拡大の推定図.×印は間を埋める分布記録.灰色は標高100m以上の場所を示す.
(国土地理院タイル「自分でつくる色別標高図」に,分布地や分布前線を記入し掲載).青木(2021)を改変.
(国土地理院タイル「自分でつくる色別標高図」に,分布地や分布前線を記入し掲載).青木(2021)を改変.
分布拡大速度については,大阪湾周辺の記録をもとに直線距離で求めてみたところ,陸続きの場合だいたい6~9km/年,途中に海峡などが入ると3~4km/年になることがわかりました(Aoki,1997).複数の分布拡大ルートで計算してもあまり大きなばらつきはありません.本種は,羽化後,羽化地を離れてある程度移動するといわれています(石田ら,1988).残念ながらどれくらい移動するかは具体的には調べられていません.しかし成虫がその一生の間(とくに羽化後)に平均して6~9kmくらいの距離を移動することは十分に考えられます.後述するように,化性は二年一化または一年一化と考えられています.したがって,二年一化でも平均してその倍の距離を移動すればこの速度を達成できることになり,上記の分布拡大は,成虫の通常の生活史の移動範囲内で行われている分散によると考えることに無理はありません.
また分布拡大の特徴として次のような事実もあります.例えば兵庫県本州側南部では,1987年に明石公園で1頭のオスが発見されたあと,1990年にそこから少し離れた垂水区で複数のオスが,1991年には明石公園やその北に伸びる櫨谷に並ぶ複数の池でオス12頭とメス2頭が,1992年には櫨谷で多数の個体が発見されました.さらにその後,1995年に加古川市で1頭のオスが発見され,その4年後には加古川市や高砂市で多数の個体が見られるようになりました.つまり,少数(1頭)の個体が発見され,その数年後に数が急増するという傾向が見られるのです.吉田・八木(2012)も愛知県の調査で同様の傾向を認め,「侵入した場所付近で繁殖を行い,個体数を増加させること.言わば核となる産地をつくることが分布拡大の必須条件といえる.」とまとめています.
一方で,分布拡大の原点ともいえる,鹿児島県,高知県,三重県の記録については,おそらく海洋を長距離移動してきたものでしょう.本種が海洋を移動する能力を有する証拠としては,南北大東島での記録(苅部・小浜,1980;杉村ら,1999)をあげることができます.南北大東島は沖縄本島から400kmほども離れており,一度も陸続きになったことがない海洋島です.人為的移入の可能性は皆無ではありませんが,これについてはむしろその証拠がないわけですからここでは考えないようにします.
以上のことを重ね合わせると,通常は「生活史範囲内の移動」と「移動先での繁殖」を繰り返しながら分布拡大していると考えることができます.しかし時には,生息可能な水域の有無や地形などによって,かなり長距離を一気に移動することもあれば,なかなか移動しない(できない)こともあって,こういったことが入り交じりながら分布を拡大しているのが実態でしょう.
また分布拡大の特徴として次のような事実もあります.例えば兵庫県本州側南部では,1987年に明石公園で1頭のオスが発見されたあと,1990年にそこから少し離れた垂水区で複数のオスが,1991年には明石公園やその北に伸びる櫨谷に並ぶ複数の池でオス12頭とメス2頭が,1992年には櫨谷で多数の個体が発見されました.さらにその後,1995年に加古川市で1頭のオスが発見され,その4年後には加古川市や高砂市で多数の個体が見られるようになりました.つまり,少数(1頭)の個体が発見され,その数年後に数が急増するという傾向が見られるのです.吉田・八木(2012)も愛知県の調査で同様の傾向を認め,「侵入した場所付近で繁殖を行い,個体数を増加させること.言わば核となる産地をつくることが分布拡大の必須条件といえる.」とまとめています.
一方で,分布拡大の原点ともいえる,鹿児島県,高知県,三重県の記録については,おそらく海洋を長距離移動してきたものでしょう.本種が海洋を移動する能力を有する証拠としては,南北大東島での記録(苅部・小浜,1980;杉村ら,1999)をあげることができます.南北大東島は沖縄本島から400kmほども離れており,一度も陸続きになったことがない海洋島です.人為的移入の可能性は皆無ではありませんが,これについてはむしろその証拠がないわけですからここでは考えないようにします.
以上のことを重ね合わせると,通常は「生活史範囲内の移動」と「移動先での繁殖」を繰り返しながら分布拡大していると考えることができます.しかし時には,生息可能な水域の有無や地形などによって,かなり長距離を一気に移動することもあれば,なかなか移動しない(できない)こともあって,こういったことが入り交じりながら分布を拡大しているのが実態でしょう.
温暖化? 北上するタイワンウチワヤンマ 改訂版
分布拡大にともなう生活史の調整
一般に温帯性の昆虫は,耐寒性のあるステージで冬を迎えることができるように,生活史をその地域(緯度といってもよい)の季節変化に同調させています.このことは,例えば分布を北に広げていくとき,その地域の季節変化に合うように生活史を調整する必要があることを意味します.タイワンウチワヤンマは,気温や日長の異なる,非常に広い緯度範囲にまたがって分布している種です.したがって,その生活史は南方と北限の日本では,かなり異なったものになっていると想像できます.
Ⅰ.タイワンウチワヤンマの生活史
残念ながら,タイワンウチワヤンマの生活史について総合的に調べられた研究報告はありません.しかし,通年にわたる非常に綿密な羽化調査が沖縄県那覇市(長嶺,1964)と高知県南国市(松本,2001)で行われています.また神戸市西区櫨谷町近辺で3年47回にわたる幼虫・羽化殻・成虫の採集結果から,生活史を推定した報告があります(Aoki, 1997).それ以外にも,わずか2頭ですが,沖縄県で採集された亜終齢幼虫の羽化までの飼育記録があります(尾花ら,1979).これらから,まずはタイワンウチワヤンマの生活史を推定してみるところから始めましょう.
Ⅰ.タイワンウチワヤンマの生活史
残念ながら,タイワンウチワヤンマの生活史について総合的に調べられた研究報告はありません.しかし,通年にわたる非常に綿密な羽化調査が沖縄県那覇市(長嶺,1964)と高知県南国市(松本,2001)で行われています.また神戸市西区櫨谷町近辺で3年47回にわたる幼虫・羽化殻・成虫の採集結果から,生活史を推定した報告があります(Aoki, 1997).それ以外にも,わずか2頭ですが,沖縄県で採集された亜終齢幼虫の羽化までの飼育記録があります(尾花ら,1979).これらから,まずはタイワンウチワヤンマの生活史を推定してみるところから始めましょう.
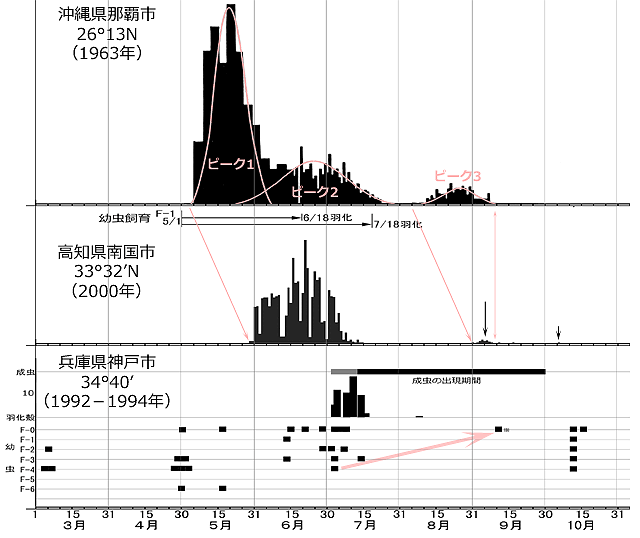
図10.タイワンウチワヤンマの生活史解明に関する情報を集約したもの.
沖縄県那覇市のグラフは長嶺(1964)にピークの曲線を追記,その下の幼虫飼育結果は尾花ら(1979)のデータを図示した.幼虫飼育は5月1日に沖縄で採集された亜終齢幼虫(F-1)を,大阪(おそらく室内)で行ったもので,羽化までの時間が少し長引いている可能性がある.高知県南国市のグラフは松本(2001)をもとにして描画した.神戸市の図はAoki(1997) Table1 掲載の,3年47日の間に得られたデータを集約して描画した.成虫の出現期間は最も早いものと遅いものの範囲を示している.※印の終齢は,採集時の所見から脱皮直後であって,その直前まで亜終齢であったことを示す.
沖縄県那覇市のグラフは長嶺(1964)にピークの曲線を追記,その下の幼虫飼育結果は尾花ら(1979)のデータを図示した.幼虫飼育は5月1日に沖縄で採集された亜終齢幼虫(F-1)を,大阪(おそらく室内)で行ったもので,羽化までの時間が少し長引いている可能性がある.高知県南国市のグラフは松本(2001)をもとにして描画した.神戸市の図はAoki(1997) Table1 掲載の,3年47日の間に得られたデータを集約して描画した.成虫の出現期間は最も早いものと遅いものの範囲を示している.※印の終齢は,採集時の所見から脱皮直後であって,その直前まで亜終齢であったことを示す.
まず神戸市のデータを見てみましょう.5月の幼虫データで越冬後の幼虫の齢範囲がF-0~F-6※2であることから,越冬している幼虫は少なくともF-0~F-6齢以下であることが分かります.そしてこの越冬した幼虫すべては,その年内には羽化しないようです.それは,羽化時期の7月中旬にF-3齢が,そして成虫の飛んでいる9月初めにF-1齢が存在し(図10のキャプション参照),さらにこのF-1齢はその年に産下された卵から育ったものとは考えられないからです(もしそうだとするとウスバキトンボなみの速さです).つまり赤矢印で示したように発育していると考えるのが妥当です.
8月の幼虫データがないので若干の不安は残りますが,成虫が飛ぶシーズンにも幼虫が途切れずにいることになり,少なくとも1世代に2年以上かかることが分かります.化性に関しては,吉田ら(2017)が愛知県の調査で,「2014年10月17日に同池に寄った際,池の水は抜かれて工事が行われていた.水が張られたのは2015年で,2016年8月12日の調査において羽化殻1オスを採集できたので,野外での幼虫期は1年」と記しています.このことから考えると,神戸における幼虫期が3年以上ということは考えにくく,基本的な化性を二年一化とするのが妥当でしょう.
8月の幼虫データがないので若干の不安は残りますが,成虫が飛ぶシーズンにも幼虫が途切れずにいることになり,少なくとも1世代に2年以上かかることが分かります.化性に関しては,吉田ら(2017)が愛知県の調査で,「2014年10月17日に同池に寄った際,池の水は抜かれて工事が行われていた.水が張られたのは2015年で,2016年8月12日の調査において羽化殻1オスを採集できたので,野外での幼虫期は1年」と記しています.このことから考えると,神戸における幼虫期が3年以上ということは考えにくく,基本的な化性を二年一化とするのが妥当でしょう.
※2.F はFinalを表し,終齢から逆に齢期を数える方法である.F-0は終齢,F-1は亜終齢,F-2は終齢二つ手前,というように数える.
次に沖縄県の羽化曲線を見ると,5月から7月にかけて連続する羽化と8月中盤から始まる羽化の,独立二峰性のパターンを示しています.こういった独立二峰性の羽化は,基本として二年一化の生活史を有するトンボの場合,その一部が分かれて一年一化で羽化する場合によく見られるパターンです.これを同時出生集団の分割(cohort splitting)といい,トンボではよく知られている現象です※3.これは吉田ら(2017)の一年一化との報告とも合致します.
沖縄県の羽化曲線と幼虫飼育結果をもとに考えると,二年一化群のピーク1は前年の冬を終齢で過ごした幼虫が羽化したもの,重なるように続くピーク2は前年の冬を亜終齢あるいはそれ以下で過ごした幼虫が羽化したものと結論できます.ピーク3は一年一化群で,さらに若齢で冬を越した幼虫が急速に成長し羽化したものと考えられます.このように羽化直前の越冬齢期にかなりの幅があることから,本種は夏季種に位置づけられます(Corbet, 1962).
沖縄県の羽化曲線と幼虫飼育結果をもとに考えると,二年一化群のピーク1は前年の冬を終齢で過ごした幼虫が羽化したもの,重なるように続くピーク2は前年の冬を亜終齢あるいはそれ以下で過ごした幼虫が羽化したものと結論できます.ピーク3は一年一化群で,さらに若齢で冬を越した幼虫が急速に成長し羽化したものと考えられます.このように羽化直前の越冬齢期にかなりの幅があることから,本種は夏季種に位置づけられます(Corbet, 1962).
※3.同時出生集団の分割に関しては,本サイトの別の記事を参照のこと.
温帯の夏季種幼虫は,そのほとんどの齢期において温度依存的に成長していると考えて間違いないでしょう(cf. Corbet, 1999:Table 7-3).ただ,沖縄県と高知県のグラフから,後半の小さな羽化ピークの終わりがほとんど同じ時期になっている点は興味深いところです.幼虫越冬するトンボにおいて耐寒性がないステージは,一般に成虫,卵,幼虫の初期数齢,アポリシス※4の進行中であるといわれています(Norling, 1984).羽化した後には,成虫の前生殖期・産卵,卵発生,孵化,若齢幼虫の発育と耐寒性のないステージがある期間続くので,秋のある時期に羽化を止め,冬の到来前に幼虫が耐寒性のある齢期まで成長する時間を確保する必要があります.つまり終齢幼虫に秋遅くの羽化を抑制する何らかの制御機構(休眠)が発達している可能性があるということです※5.
以上をまとめると,日本におけるタイワンウチワヤンマの生活史は次のようにまとめられます.またそれを模式的に図示すると,図11のように描くことができます.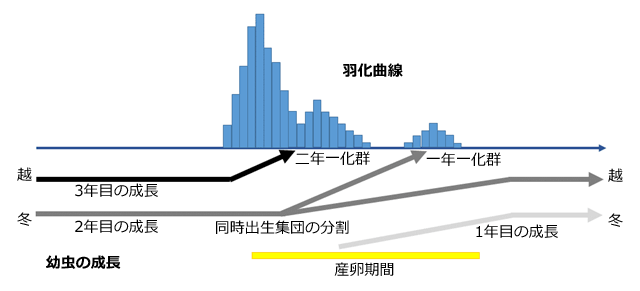
以上をまとめると,日本におけるタイワンウチワヤンマの生活史は次のようにまとめられます.またそれを模式的に図示すると,図11のように描くことができます.
- 幼虫越冬性の夏季種である.
- 化性は二年一化が基本で,一部は同時出生集団の分割により一年一化となる.
- 二年一化群はシーズン最初の大きな羽化ピークをつくり,一年一化群はシーズン後半の小さな羽化ピークをつくる.
図11.タイワンウチワヤンマの生活史(推定)を模式的に示したもの.
沖縄県の羽化曲線を例にして,日本におけるタイワンウチワヤンマの生活史を模式的に示したもの.高知県においても,小さな秋口の羽化が見られるので,基本的に同じであるものと思われる.同時出生集団の分割の時期(つまり幼虫の運命が一年一化の方向になるものと二年一化の方向になるものとが別れる時期)については図に示した時期とは限らない.図はどこかで起きるということを示していると考えてほしい.
沖縄県の羽化曲線を例にして,日本におけるタイワンウチワヤンマの生活史を模式的に示したもの.高知県においても,小さな秋口の羽化が見られるので,基本的に同じであるものと思われる.同時出生集団の分割の時期(つまり幼虫の運命が一年一化の方向になるものと二年一化の方向になるものとが別れる時期)については図に示した時期とは限らない.図はどこかで起きるということを示していると考えてほしい.
※4.アポリシスとは,簡単に言えば,脱皮が始まる前に,旧い表皮から内表面が剥離し,新しい表皮がつくられる過程を指す.
※5.Corbet(1999):Table 7-3の生活史分類で,B.2.2.1に該当する制御機構.B.2.2.1とは,化性は条件的で多化性,年一化性,または多年一化性,通常最後の1齢あるいはそれより前の齢期の幼虫(夏,冬)に,...,条件的休眠があって,幼虫期は1~6年あるいはそれ以上,と定義されている.
Ⅱ.北上にともなう生活史の変化
それでは次に,北上にともなう生活史の変化を見ていくことにしましょう.前提として,前節で考察したタイワンウチワヤンマの生活史が正しいものとしておきます.
まずはっきりと分かることは,羽化の開始時期が北上するにしたがって遅れているということです(図10).冬の間低温で発育が休止していた幼虫は,春になり発育零点を超えると発育を再開します.羽化前の成長は,有効積算温度の法則にしたがう温度依存の過程と考えて差しつかえないでしょう.また羽化そのものには閾値があることが知られています(Corbet, 1999).そこで石垣島,沖縄島,高知市,神戸市の4地点で,羽化が始まる時期の気温(前後10日間の平均値)を調べてみました(図12).

それでは次に,北上にともなう生活史の変化を見ていくことにしましょう.前提として,前節で考察したタイワンウチワヤンマの生活史が正しいものとしておきます.
まずはっきりと分かることは,羽化の開始時期が北上するにしたがって遅れているということです(図10).冬の間低温で発育が休止していた幼虫は,春になり発育零点を超えると発育を再開します.羽化前の成長は,有効積算温度の法則にしたがう温度依存の過程と考えて差しつかえないでしょう.また羽化そのものには閾値があることが知られています(Corbet, 1999).そこで石垣島,沖縄島,高知市,神戸市の4地点で,羽化が始まる時期の気温(前後10日間の平均値)を調べてみました(図12).
図12.羽化開始時期の緯度による違いとその時の旬(10日間)平均気温(最寄りの気象台).
実際は水温で評価されるべきものであるが,気温と水温に相関があると前提している.
神戸市の1993年の羽化時期はちょうど旬の境目に位置したため,旬平均気温は2旬を平均したものである.没姿期のはっきりしていないもの(枠を点線で表示)は,データがないことを示す.上段の数字は月を示す.1)石垣島の羽化時期・没姿期は渡辺・小浜(1985)から得た.2)沖縄島の羽化時期は長嶺(1964)から得た.3)中村市の羽化時期・没姿期は杉村ら(1989)から得た.4)神戸市のデータは筆者の調査によるもの.
図12から分かるように,本種の羽化は旬平均気温が22~23℃台になった頃に始まっていて,当然のことながら,北の方が時期的に遅くなっています.神戸市における羽化開始が,記録的な冷夏であった1993年より,異常猛暑であった1994年の方が約15日も早くなっています.このように気象条件が大きく違っても羽化する時期の平均気温は大きくは変化していません.もちろん何事にも例外的なことはあって,早い時期の羽化として,1994年4月30日鹿児島県川内市(松木ら,1994)というのがあります(1994年のこの前後の川内市旬平均は18.6℃です).一方当然のことながら,没姿期は成虫の寿命によってまちまちになっています.このように,羽化開始に関しては,温度に対するタイワンウチワヤンマの反応は,北上してもほとんど変化していないということが分かります.実際は水温で評価されるべきものであるが,気温と水温に相関があると前提している.
神戸市の1993年の羽化時期はちょうど旬の境目に位置したため,旬平均気温は2旬を平均したものである.没姿期のはっきりしていないもの(枠を点線で表示)は,データがないことを示す.上段の数字は月を示す.1)石垣島の羽化時期・没姿期は渡辺・小浜(1985)から得た.2)沖縄島の羽化時期は長嶺(1964)から得た.3)中村市の羽化時期・没姿期は杉村ら(1989)から得た.4)神戸市のデータは筆者の調査によるもの.
一方で幼虫成長に関しては,一般に北の方ほど発育零点を超える日数が少なくなって年間の有効積算温度が小さくなるはずですから,幼虫の成長が遅れ,幼虫期が長くなると予想されます.それは図10にも現れています.高知県の一年一化群の羽化ピークが沖縄県に比べて小さいことや,高知県で沖縄県のピーク1に該当するピークがはっきりと現れていないことは,いずれも全体として幼虫成長が遅くなっていること(幼虫期間の長期化)を示しているといえます※6.このことから考えると,北へ行くほど,一年一化の個体数が減少し,二年一化の生活史を送るものが大部分になるように化性が変化していくのではないかと思われます.
反対に,南の分布地域では一年一化の個体数が増加することが予想されます.これに関してはほとんど情報がありませんが,一つの興味深いデータを紹介しておきましょう.図13は,ベトナムやラオスで採集されたタイワンウチワヤンマのオス成虫と,神戸市で採集されたオス成虫の,腹長と後翅長を調べプロットしたものです.これから明らかなように,低緯度の個体のサイズが有意に小さくなっています.
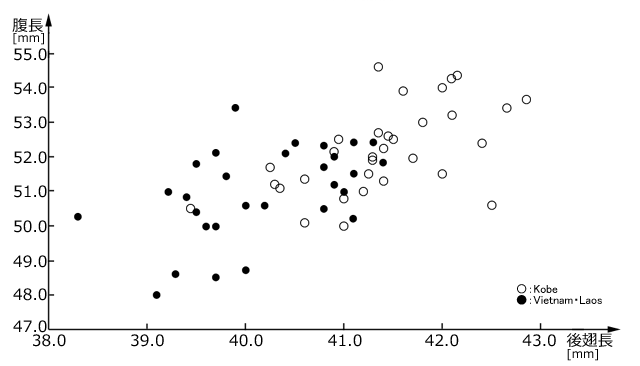
反対に,南の分布地域では一年一化の個体数が増加することが予想されます.これに関してはほとんど情報がありませんが,一つの興味深いデータを紹介しておきましょう.図13は,ベトナムやラオスで採集されたタイワンウチワヤンマのオス成虫と,神戸市で採集されたオス成虫の,腹長と後翅長を調べプロットしたものです.これから明らかなように,低緯度の個体のサイズが有意に小さくなっています.
図13.ベトナム・ラオスと神戸の個体(オスのみ)のサイズの違い.
●:ベトナム・ラオス,○:神戸.平均値±標準誤差は,腹長がそれぞれ51.0±0.23mm,52.2±0.22mm,後翅長がそれぞれ40.2±0.14mm,41.4±0.13mmであって,いずれも有意の差がある(p<0.01).ベトナム・ラオスの標本に関しては片谷(2017)の記載の採品について著者より計測値を私信にていただいたもの.神戸の標本は筆者のもの.
●:ベトナム・ラオス,○:神戸.平均値±標準誤差は,腹長がそれぞれ51.0±0.23mm,52.2±0.22mm,後翅長がそれぞれ40.2±0.14mm,41.4±0.13mmであって,いずれも有意の差がある(p<0.01).ベトナム・ラオスの標本に関しては片谷(2017)の記載の採品について著者より計測値を私信にていただいたもの.神戸の標本は筆者のもの.
※6.高緯度で幼虫の成長できる季節が短くなると幼虫の発育完了が遅れ,一年一化で羽化できる個体が少なくなったり,二年目の越冬時期に羽化直前の終齢幼虫にまで成長できる個体も少なくなるであろう.したがって,一年一化の羽化ピークが小さくなり,越冬後の羽化開始直後の大きな羽化ピークも見られなくなるということ.
幼虫成長は,温度環境に影響を受けることがあるといわれています.それに関してよく知られているのが,温帯地域の一年二化するトンボの成虫サイズで,二化目の方が体サイズが小さい傾向にあるというものです※7.Corbet(1999)はこれを少し拡張して,「羽化するトンボの成虫サイズは,一般に季節進行とともに小さくなる傾向がある」とし,その理由として,「幼虫は高温の方が速く成長するがそれと引き換えにその成長比※8を小さくするため」だと考えています.一年二化するトンボにおいては,春早くに羽化する一化目は前年の秋から冬にかけての低温期に幼虫が成長しており,夏や秋口に羽化する二化目は春から夏の高温期に幼虫が成長していますので,二化目の方がサイズが小さくなるという解釈になります.
もちろんタイワンウチワヤンマは一年二化ではありませんが,幼虫期により高温に長期間さらされるベトナムやラオスの個体の成長速度が高くかつ成長比が小さくなって,その結果成虫の体サイズが小さくなったのかも知れません.逆に考えれば,神戸では,幼虫期により低温で発育する時間が長いので成長速度が遅くかつ成長比が大きくなるので,成虫の体サイズが大きいという理屈になります.成長速度が遅いことは化性の減少につながりますから,図13は化性の変化に関係していると見ることができます.
さて,先に述べたように,幼虫越冬するトンボでは卵発生や若齢幼虫の時期に耐寒性がありません.したがって,北上するにともなって,羽化の早い二年一化群が増え,遅い一年一化群がなくなっていくということは,冬の到達の前に耐寒性のある齢期にまで成長できることにつながり,高緯度への分布拡大にとって適応的です(青木,2021).
Gurney et al.(1992)は,このような単峰性・二峰性の羽化変化(つまりこの場合二年一化と一年一化の二峰性の羽化から二年一化だけの単峰性羽化へと変化すること)は,広い緯度範囲に最適な生活史を持つ幼虫越冬種によく見られることを,数理生物学の手法を使って説明しています.またCorbet(1999)はこれを条件的化性とよび,この能力は,幼虫越冬するトンボが広い緯度範囲に分布するための必要条件であると述べています.タイワンウチワヤンマはこの条件にあてはまり,北上にともなう生活史の調整能力を有しているといえるでしょう(青木,2021).
もちろんタイワンウチワヤンマは一年二化ではありませんが,幼虫期により高温に長期間さらされるベトナムやラオスの個体の成長速度が高くかつ成長比が小さくなって,その結果成虫の体サイズが小さくなったのかも知れません.逆に考えれば,神戸では,幼虫期により低温で発育する時間が長いので成長速度が遅くかつ成長比が大きくなるので,成虫の体サイズが大きいという理屈になります.成長速度が遅いことは化性の減少につながりますから,図13は化性の変化に関係していると見ることができます.
さて,先に述べたように,幼虫越冬するトンボでは卵発生や若齢幼虫の時期に耐寒性がありません.したがって,北上するにともなって,羽化の早い二年一化群が増え,遅い一年一化群がなくなっていくということは,冬の到達の前に耐寒性のある齢期にまで成長できることにつながり,高緯度への分布拡大にとって適応的です(青木,2021).
Gurney et al.(1992)は,このような単峰性・二峰性の羽化変化(つまりこの場合二年一化と一年一化の二峰性の羽化から二年一化だけの単峰性羽化へと変化すること)は,広い緯度範囲に最適な生活史を持つ幼虫越冬種によく見られることを,数理生物学の手法を使って説明しています.またCorbet(1999)はこれを条件的化性とよび,この能力は,幼虫越冬するトンボが広い緯度範囲に分布するための必要条件であると述べています.タイワンウチワヤンマはこの条件にあてはまり,北上にともなう生活史の調整能力を有しているといえるでしょう(青木,2021).
以上,高緯度の個体群は,羽化開始の温度に対する反応が変わっていないこと,有効積算温度の減少については化性を変化させることで対応していることなどから,タイワンウチワヤンマは,本来持っている温度環境に対する生理的反応性を大きく変化させることなく,北上していると考えられます※9.つまり,生活史調整の必要性は北上を妨げる要因にはなっていないといえるでしょう.
※9.唯一調整の必要があるとすれば,秋の羽化終了を決める環境合図に対する反応性だろう(図10参照).これについては,遅く羽化した個体やその子孫が冬期の低温で淘汰されることで個体群が小進化し,適応性を獲得していっているのかもしれない.
温暖化? 北上するタイワンウチワヤンマ 改訂版
分布拡大と種間競争の問題
新しい種がそれまで分布していなかった地域に進入すると,ニッチの近い先住種との間に種間競争が起きる可能性があります.よく引き合いに出されているのがウチワヤンマとの競争です.ウチワヤンマとタイワンウチワヤンマは,ともにため池などの池沼性生活者であり,出現季節も大きく重なっています.オス成虫は,いずれも池の畔に突き立った棒や茎などに静止して縄張りを形成します.メス成虫は,通常水面に浮かぶ浮遊物等に腹端を打ちつけて糸状の構造物の着いた卵を絡ませるように産卵します.もっとも,タイワンウチワヤンマは,水面を飛び回って打水産卵することもあるようです.

写真4.タイワンウチワヤンマ(左)とウチワヤンマ(右)のオスの静止.
ともに池の畔の棒や茎の先などに止まって縄張りを形成する.左:2014.7.28. 南あわじ市,右:2020.7.19. 大阪府.
ともに池の畔の棒や茎の先などに止まって縄張りを形成する.左:2014.7.28. 南あわじ市,右:2020.7.19. 大阪府.

写真5.タイワンウチワヤンマ(左)とウチワヤンマ(右)のメスの産卵行動.
ともに池に岸近くなどの浮遊物に腹端を打ちつけて卵をからめるようにして産卵する.腹端を打ちつけるポイントをねらって,水面すぐ上でホバリングしている.左:2021.8.1. 南あわじ市,右:2021.6.25. 加東市.
ともに池に岸近くなどの浮遊物に腹端を打ちつけて卵をからめるようにして産卵する.腹端を打ちつけるポイントをねらって,水面すぐ上でホバリングしている.左:2021.8.1. 南あわじ市,右:2021.6.25. 加東市.
しかし幼虫の生活場所は両種でかなり異なっているようです.タイワンウチワヤンマ幼虫は,比較的浅いところで見つかることが多く,ウチワヤンマ幼虫は池の深いところで生活していて,採集時に一緒に網に入るようなことはまずありません.ただし,タイワンウチワヤンマ幼虫は冬季に移動するようで,図10の神戸市での調査時,1992年12月29日,1993年2月13日,25日,3月18日,27日,4月12日と続けて,それまで採れた場所で幼虫がまったく採集されませんでした.おそらく深みに移動しているのでしょう.ただ羽化については類似の場所で行われることもあり,同日・同所に羽化殻が付いていたのを観察しています.

写真6.タイワンウチワヤンマ(左)とウチワヤンマ(右)の羽化殻が同じ日の同じ植生の場所に着いている.2020.7.12. 加東市.
両種の生息する池沼環境には,かなりの重なりがあるようです.1991年から兵庫県南部の播磨平野に分布が拡大し2020年で30年経ちましたが,先住種のウチワヤンマのいた152カ所の池の91カ所(約60%)にタイワンウチワヤンマが入りこみました(青木,2021).しかし一方で,ウチワヤンマしか見られない池や,タイワンウチワヤンマしか見られない池もあり,微妙に好みとする池沼環境が異なっている印象もあります.その場合,ウチワヤンマは植生が少なくより大規模な池に,タイワンウチワヤンマは植生が密に茂った比較的小規模の池に見られることが多いようです.

写真7.タイワンウチワヤンマ(左)とウチワヤンマ(右)の生息環境の違いの典型例.
タイワンウチワヤンマは比較的植生が密に茂った池にもよく入りこんで活動している.一方ウチワヤンマは植生がなく開水面が広がる大規模な池によく入りこんでいる.左:2009.8.14. 南あわじ市.右:2010.8.8. 稲美町.
タイワンウチワヤンマは比較的植生が密に茂った池にもよく入りこんで活動している.一方ウチワヤンマは植生がなく開水面が広がる大規模な池によく入りこんでいる.左:2009.8.14. 南あわじ市.右:2010.8.8. 稲美町.
このように,両種はニッチが完全に重なっているわけではありませんが,それでも成虫時代には競争が生じているようで,いくつかの観察例が報告されています.成田(1988)は,「ウチワヤンマの縄張り域にタイワンウチワヤンマが侵入しても追うことをせず,筆者がウチワヤンマを捕虫網の柄で枝をゆすり飛ばしてやると,近くにいたタイワンウチワヤンマに次々に追われてしまった」と記しており,平井(1980)は,「1968年徳島市では,(タイワンウチワヤンマが)溜池・休耕田・水田間の小川等いたるところに普通に見られ,在来のウチワヤンマが,侵入者のタイワンウチワヤンマに駆逐されたのではないかと思われるほど著しく減少した」と記しています.
そのほかにもありますが,大体はタイワンウチワヤンマの方が競争に強いような記述です.一方で競争の結果でしょうか,ニッチの分化が起きているような記述も見られます.成田(1988)は,「両種の間にはウチワヤンマの方が出現が早く,棲み分けが行われているように考えられる」と述べており,吉田・八木(2012)は,「沿岸部に近い地域にはタイワンウチワヤンマが増え,ウチワヤンマは内陸部に追いやられている傾向が強い」と述べています.
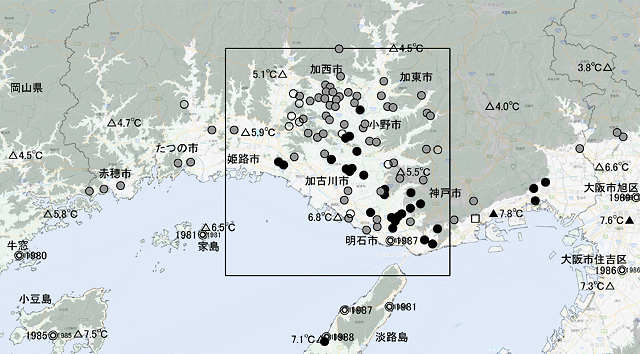
兵庫県の播磨地域では,1991年頃からタイワンウチワヤンマの分布が広がり,2020年現在では,東西は赤穂市から神戸市に,南北は加西・加東市から明石市に至る,西・中・北・東播磨全域に分布しています(図14).
まず使用するデータは図14の長方形で囲まれた範囲にしました.この範囲は1991年から2020年までの30年間順次分布が拡大していて,分布拡大にともなう10年ごとの変化がうまく現れるだろうと考えたからです.データにはほとんど3次メッシュコードが記載されており,このメッシュ区画を調査場所の単位としました.範囲内の全メッシュ区画のうち,解析対象としたのは,タイワンウチワヤンマおよびウチワヤンマのどちらか一方あるいは両方が見つかったところだけにしました.そしてある日付にあるメッシュ区画を訪れたときに,1回の調査が行われたとしました.同日に2回同じメッシュ区画を訪れたことはありません.そしてある旬(10日間)の10年間の全観察個体数を全調査数で割ったものをその10年のその旬の個体数としました.ただし観察個体数は時に極端に大きな数値が記録されることがあり,これによる影響を軽減するために,10個体以上は10個体として計算しています.
以上をまとめたのが図15です.
そのほかにもありますが,大体はタイワンウチワヤンマの方が競争に強いような記述です.一方で競争の結果でしょうか,ニッチの分化が起きているような記述も見られます.成田(1988)は,「両種の間にはウチワヤンマの方が出現が早く,棲み分けが行われているように考えられる」と述べており,吉田・八木(2012)は,「沿岸部に近い地域にはタイワンウチワヤンマが増え,ウチワヤンマは内陸部に追いやられている傾向が強い」と述べています.
兵庫県の播磨地域では,1991年頃からタイワンウチワヤンマの分布が広がり,2020年現在では,東西は赤穂市から神戸市に,南北は加西・加東市から明石市に至る,西・中・北・東播磨全域に分布しています(図14).
図14.播磨地域におけるタイワンウチワヤンマの分布拡大の様相.1987年の明石公園の発見以後,2010年頃にはほぼ播磨地域全域に分布が広がった.(青木,2021原図)
◎は1980年代,●は1991-2000年,●は2001-2010年,○は2011-2020年それぞれに初記録が出た場所を示す.△,▲はそれぞれアメダスおよび地方気象台設置場所を示し,その横の数値は2010~2021年までの冬季(12月~2月)の平均気温を示す.長方形で囲まれた地域は,成虫の活動時期を調べた地域の範囲.灰色で塗られた部分は標高100m以上の場所を示す.
国土地理院タイル「自分でつくる色別標高図」に記録場所などを記入し掲載.
この地域では,最近ウチワヤンマの活動時期が以前より早くなっているように感じられます.地元の研究家も同様のことを述べています.そこで,タイワンウチワヤンマとウチワヤンマ成虫の季節消長について,観察データをもとに調べてみました.単に出現時期の範囲だけでなく,10年ごとの季節進行にともなう個体数の変化を見るためのデータ処理を行いました.◎は1980年代,●は1991-2000年,●は2001-2010年,○は2011-2020年それぞれに初記録が出た場所を示す.△,▲はそれぞれアメダスおよび地方気象台設置場所を示し,その横の数値は2010~2021年までの冬季(12月~2月)の平均気温を示す.長方形で囲まれた地域は,成虫の活動時期を調べた地域の範囲.灰色で塗られた部分は標高100m以上の場所を示す.
国土地理院タイル「自分でつくる色別標高図」に記録場所などを記入し掲載.
まず使用するデータは図14の長方形で囲まれた範囲にしました.この範囲は1991年から2020年までの30年間順次分布が拡大していて,分布拡大にともなう10年ごとの変化がうまく現れるだろうと考えたからです.データにはほとんど3次メッシュコードが記載されており,このメッシュ区画を調査場所の単位としました.範囲内の全メッシュ区画のうち,解析対象としたのは,タイワンウチワヤンマおよびウチワヤンマのどちらか一方あるいは両方が見つかったところだけにしました.そしてある日付にあるメッシュ区画を訪れたときに,1回の調査が行われたとしました.同日に2回同じメッシュ区画を訪れたことはありません.そしてある旬(10日間)の10年間の全観察個体数を全調査数で割ったものをその10年のその旬の個体数としました.ただし観察個体数は時に極端に大きな数値が記録されることがあり,これによる影響を軽減するために,10個体以上は10個体として計算しています.
以上をまとめたのが図15です.
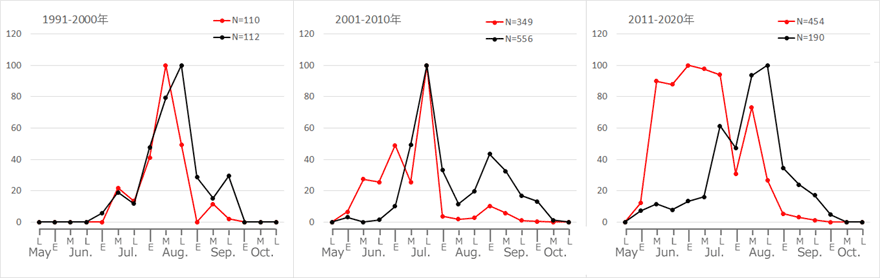
図15.タイワンウチワヤンマとウチワヤンマの出現時期の推移.赤色:ウチワヤンマ,黒色:タイワンウチワヤンマ.
E,M,Lはそれぞれ上旬,中旬,下旬を表す.グラフは両者の消長を比較しやすくするため,最大個体数を100とする相対値で描いている.青木(2021)を改変.
E,M,Lはそれぞれ上旬,中旬,下旬を表す.グラフは両者の消長を比較しやすくするため,最大個体数を100とする相対値で描いている.青木(2021)を改変.
タイワンウチワヤンマが播磨地域に出現して分布を拡大し始めた1991年からの10年間は,両種ともほぼ同じような季節消長を示しています.しかし,播磨地域全域に分布が拡大していった2001年以降,次第にウチワヤンマが早く出現するように変化しています.一方でタイワンウチワヤンマの方は,若干出現ピーク時期のゆらぎ※10はありますが,ウチワヤンマほど大きな変化はありません.この30年間で,両種ニッチの時間的な分化が生じたことが分かります.
この時間的なニッチ分化を引き起こした原因については,まず両種の種間競争が頭に浮かびますが,最近の気温上昇(気温変化については後に示す)がこのようなニッチ分化を引き起こしたと考えることも可能です.実際グラフをよく見ると,タイワンウチワヤンマ成虫の出現開始時期も,この30年の間に7月上旬から6月上旬へと,約1ヶ月早まっています.ウチワヤンマほど極端ではありませんが,タイワンウチワヤンマにも出現時期の早期化が見られるのです.そこでさらに,ウチワヤンマやタイワンウチワヤンマと同時期に同所的に活動している8種のトンボに,少し生息環境が異なるオオシオカラトンボを加えて,解析方法をほぼ同じ(1990年以前のデータを含めた点が異なる)にして消長変化を調べてみました.図16がその結果です.
この時間的なニッチ分化を引き起こした原因については,まず両種の種間競争が頭に浮かびますが,最近の気温上昇(気温変化については後に示す)がこのようなニッチ分化を引き起こしたと考えることも可能です.実際グラフをよく見ると,タイワンウチワヤンマ成虫の出現開始時期も,この30年の間に7月上旬から6月上旬へと,約1ヶ月早まっています.ウチワヤンマほど極端ではありませんが,タイワンウチワヤンマにも出現時期の早期化が見られるのです.そこでさらに,ウチワヤンマやタイワンウチワヤンマと同時期に同所的に活動している8種のトンボに,少し生息環境が異なるオオシオカラトンボを加えて,解析方法をほぼ同じ(1990年以前のデータを含めた点が異なる)にして消長変化を調べてみました.図16がその結果です.
※10.このゆらぎの存在は興味深い現象といえる.北上してきたタイワンウチワヤンマが,有効温量の減少によって幼虫成長速度が遅れ,羽化のタイミングが変わってくる状況を調整しつつあることの表れなのかも知れない.
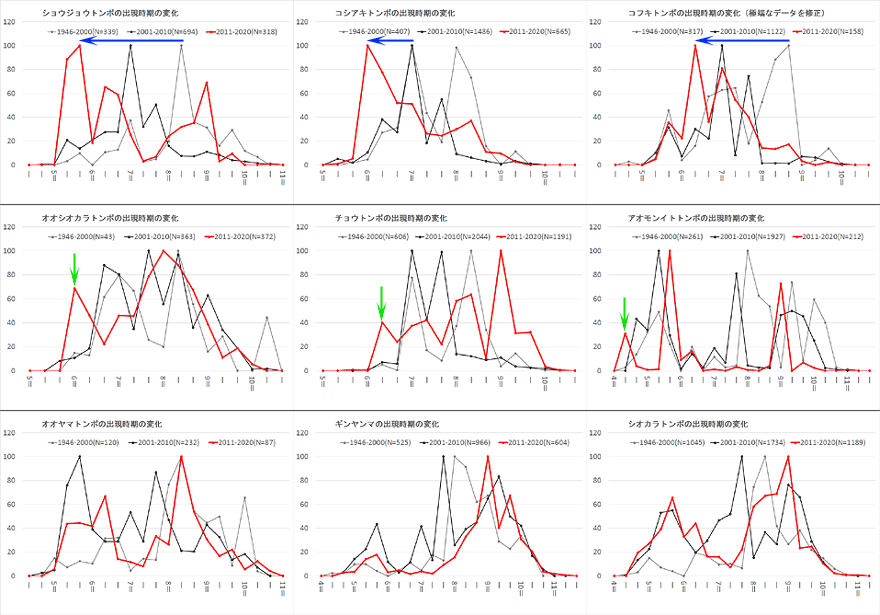
図16.同所的9種の季節消長の変化.灰色:1946-2000年,黒色:2001-2010年,赤色:2011-2020年.ずっと以前からいる種たちなので,より傾向をはっきりと出すために1990年以前のデータを加えている.
ショウジョウトンボ,コシアキトンボ,コフキトンボに,青矢印で示したように出現ピークの早期シフトが顕著に見られる.オオシオカラトンボ,チョウトンボ,アオモンイトトンボにはそういった早期化シフトは見られないが,出現時期の初めにやや小さなピーク(緑矢印)が現れている.オオヤマトンボ,ギンヤンマ,シオカラトンボにはそれらの兆候は見られない.
ショウジョウトンボ,コシアキトンボ,コフキトンボに,青矢印で示したように出現ピークの早期シフトが顕著に見られる.オオシオカラトンボ,チョウトンボ,アオモンイトトンボにはそういった早期化シフトは見られないが,出現時期の初めにやや小さなピーク(緑矢印)が現れている.オオヤマトンボ,ギンヤンマ,シオカラトンボにはそれらの兆候は見られない.
これを見ると,この10年間(赤色),ショウジョウトンボ,コシアキトンボ,コフキトンボに,青矢印で示したような出現ピークの早期化がはっきりと現れています※11.またオオシオカラトンボ,チョウトンボ,アオモンイトトンボにも,前3種ほどはっきりとはしませんが,早期に出現する個体の増加傾向がうかがえます(緑矢印).しかし残りの3種,オオヤマトンボ,ギンヤンマ,シオカラトンボにはそれは見られません.用いた観察データは,このような目的のために計画的に取られたものではありませんので,その正確性に問題が潜んでいる可能性はないとはいえませんが,この結果は最近の観察経験によく合っている印象はあります.
なお図16で種によって異なる結果が出たのは,幼虫の温度に対する反応が異なるためかも知れません.しかし早期化傾向が見られる種の存在は,この現象がウチワヤンマに特有のものではないことを示しています.もちろんこのデータだけでは,競争による結果であるということを否定できませんが,上述のニッチ分化は最近の気温上昇による結果であるとするのが妥当なように思えます.
このように,気温上昇の影響というのはその地域の在来種にも働くので,北上してきた南方種との種間関係について考える際には,全体として群集がどのように変化していくのかといった視点が必要です.ここでの議論においては,次のように一般化できるでしょう.
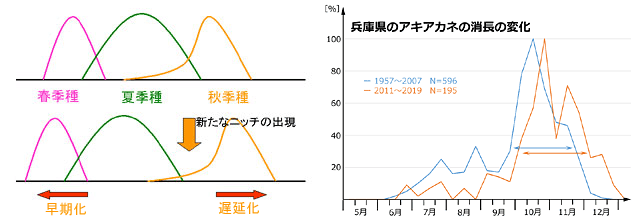
いくつかの夏季種※12は,図16に示したように成虫出現時期のピークが早い方にずれてきています.また秋季種といわれるアカネ属やアオイトトンボ属の一部には,その繁殖活動期が遅い方にずれている種がいます.アキアカネがその例で,兵庫県南部では以前は9月下旬から11月下旬ぐらいまでが繁殖活動がもっとも盛んに行われる時期でしたが,最近は10月上旬から12月上旬ぐらいまでがその時期になっています(図17右).こういった種が増えてくると,ちょうどその間にニッチの空白が生じます(図17左).
なお図16で種によって異なる結果が出たのは,幼虫の温度に対する反応が異なるためかも知れません.しかし早期化傾向が見られる種の存在は,この現象がウチワヤンマに特有のものではないことを示しています.もちろんこのデータだけでは,競争による結果であるということを否定できませんが,上述のニッチ分化は最近の気温上昇による結果であるとするのが妥当なように思えます.
このように,気温上昇の影響というのはその地域の在来種にも働くので,北上してきた南方種との種間関係について考える際には,全体として群集がどのように変化していくのかといった視点が必要です.ここでの議論においては,次のように一般化できるでしょう.
いくつかの夏季種※12は,図16に示したように成虫出現時期のピークが早い方にずれてきています.また秋季種といわれるアカネ属やアオイトトンボ属の一部には,その繁殖活動期が遅い方にずれている種がいます.アキアカネがその例で,兵庫県南部では以前は9月下旬から11月下旬ぐらいまでが繁殖活動がもっとも盛んに行われる時期でしたが,最近は10月上旬から12月上旬ぐらいまでがその時期になっています(図17右).こういった種が増えてくると,ちょうどその間にニッチの空白が生じます(図17左).
図17.左:気温上昇によって夏に新たなニッチの空白が生じることを示す模式図.右:秋季種の一種アキアカネの繁殖活動期が兵庫県南部で遅れていることを示すグラフ.
左は.気温上昇によって春季種・夏季種の出現が早期化し,秋の気温低下が遅れて秋季種の成虫活動期が遅延化すると,その間にニッチの空白が生じることを示している.夏季種では図16のコシアキトンボ,秋季種では上図右側に示したアキアカネがそれぞれの具体例となる.春季種の早期化については具体例がなく仮説である.アキアカネの消長変化のグラフは基本的に図16と同じ処理で描いているが,最近の傾向を際立たせるために移行的数値が多く見られる2008-2010年のデータをあえて省いている.
以上をまとめると,タイワンウチワヤンマはウチワヤンマ等の活動時期早期化へのシフトにより空いたニッチに入りこんで競争が軽減されていると見ることができ,種間競争は分布北上を困難にする大きな要因にはなっていないと考えられます.左は.気温上昇によって春季種・夏季種の出現が早期化し,秋の気温低下が遅れて秋季種の成虫活動期が遅延化すると,その間にニッチの空白が生じることを示している.夏季種では図16のコシアキトンボ,秋季種では上図右側に示したアキアカネがそれぞれの具体例となる.春季種の早期化については具体例がなく仮説である.アキアカネの消長変化のグラフは基本的に図16と同じ処理で描いているが,最近の傾向を際立たせるために移行的数値が多く見られる2008-2010年のデータをあえて省いている.
温暖化? 北上するタイワンウチワヤンマ 改訂版
分布拡大における温度要因の検討
さて,分布北上にともなう生活史の調整や種間競争の問題は,タイワンウチワヤンマの場合あまり問題にならないということを見てきました.では,南方種である本種がずっと昔に分布を北上させず,「今」になって北へ分布を広げてきているのはなぜでしょうか.それは,「今」起きている何かが,分布北上をはばんでいた何かを取り払ったからと考えると,その理由が見えてくるかも知れません.こういった文脈でよく引き合いに出されるのが『温暖化※13』です.ここでは,この『温暖化』と分布北上にの関係について考えてみたいと思います.
温暖化,つまり気温の上昇は生物にさまざまの影響を及ぼします.タイワンウチワヤンマについていえば,北上にともなって遅れていく幼虫の成長は,春から秋の気温上昇によって相殺され,その生活史の調整をより緩やかなものにするでしょう.また種間競争では,気温上昇が先住種の生活史に影響を与え新しいニッチを生み出す可能性にも言及しました.これら以外にもう一つ考えられることは,冬季の気温上昇が幼虫の越冬可能性を高めるかも知れないということでしょう.
現在冬の厳しい山陰地方にまで分布が拡大していることを考えると,北上にともなって幼虫が何らかの耐寒性を獲得している可能性がないとは言い切れません.また図10に表れている一年一化群の羽化終了時期が沖縄と高知でほぼ同じになっていることは,休眠機構を含めた冬を迎えるための何らかの生理的なしくみを獲得していることを予想させます.しかしこれらはすべて想像に過ぎずここでは議論できません.したがってここではこのような生理的しくみの有無にかかわらず,越冬幼虫の耐寒性は北上にともなって大きく変化していないという前提で,冬季気温の変化と分布拡大の相関性についてだけ見ていきたいと思います.
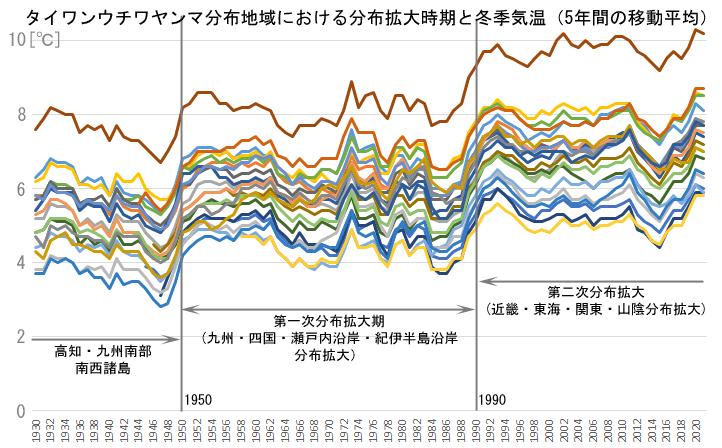
ではまずは,冬季気温が最近本当に上昇しているかどうか確かめておきましょう.図18は現在タイワンウチワヤンマが分布している地域の各気象台の,過去90年の冬季気温(12月-1月)平均値の変化を,5年間の移動平均で示したものです.これを見ると,明らかに冬季気温はこの90年間上昇傾向があるといえます.しかし冬季気温上昇は一律に起きているのではなく,1950年前後と1990年前後に急激に上昇するような変化をしています.それ以外の時期は上下しながらもほぼ同じような温度が続くように推移し,全体として階段状の上昇曲線になっています.
これだけを見ると,南方種の分布拡大が冬季気温上昇で説明できるのではないかという印象が強くなりそうです.そこで,温暖化とは異なる視点で分布拡大(北上)を解釈する議論を一つ検討しておきましょう.
温暖化,つまり気温の上昇は生物にさまざまの影響を及ぼします.タイワンウチワヤンマについていえば,北上にともなって遅れていく幼虫の成長は,春から秋の気温上昇によって相殺され,その生活史の調整をより緩やかなものにするでしょう.また種間競争では,気温上昇が先住種の生活史に影響を与え新しいニッチを生み出す可能性にも言及しました.これら以外にもう一つ考えられることは,冬季の気温上昇が幼虫の越冬可能性を高めるかも知れないということでしょう.
現在冬の厳しい山陰地方にまで分布が拡大していることを考えると,北上にともなって幼虫が何らかの耐寒性を獲得している可能性がないとは言い切れません.また図10に表れている一年一化群の羽化終了時期が沖縄と高知でほぼ同じになっていることは,休眠機構を含めた冬を迎えるための何らかの生理的なしくみを獲得していることを予想させます.しかしこれらはすべて想像に過ぎずここでは議論できません.したがってここではこのような生理的しくみの有無にかかわらず,越冬幼虫の耐寒性は北上にともなって大きく変化していないという前提で,冬季気温の変化と分布拡大の相関性についてだけ見ていきたいと思います.
ではまずは,冬季気温が最近本当に上昇しているかどうか確かめておきましょう.図18は現在タイワンウチワヤンマが分布している地域の各気象台の,過去90年の冬季気温(12月-1月)平均値の変化を,5年間の移動平均で示したものです.これを見ると,明らかに冬季気温はこの90年間上昇傾向があるといえます.しかし冬季気温上昇は一律に起きているのではなく,1950年前後と1990年前後に急激に上昇するような変化をしています.それ以外の時期は上下しながらもほぼ同じような温度が続くように推移し,全体として階段状の上昇曲線になっています.
図18.全国の分布地域の冬季平均気温変化.
アメダスは1974年から順次運用を始めていて,旧い気温データは気象台にしかないため,現在分布している地域内の気象台のデータで示している.したがってこれらの気温変化は,必ずしも生息地に近い場所での気温でないことは注意しておく必要がある.
気象台は次の通り.福岡,熊本,鹿児島,松山,徳島,高松,下関,広島,岡山,神戸,大阪,京都,和歌山,松江,鳥取,津,名古屋,静岡,横浜,東京,千葉.第二次分布拡大期で,一つの目安となる6℃(図22参照)を恒常的に下回っている地域は,京都,奈良,鳥取,松江,名古屋で,山陰,盆地など冬季に気温が下がりやすい地域である.飛び抜けて高いのは鹿児島.
この階段状のグラフで,1950~1990年の平坦期に,九州・四国・瀬戸内海・紀伊半島沿岸域の分布拡大をほぼ終え,さらにその後の近畿地方内奥部・東海・関東,そしてと山陰地方への分布拡大が,1990年以降の一段高くなった平坦期に起きています.分布拡大時期と冬季気温の上昇は,偶然かも知れませんが,よい相関を示しているように見えます.アメダスは1974年から順次運用を始めていて,旧い気温データは気象台にしかないため,現在分布している地域内の気象台のデータで示している.したがってこれらの気温変化は,必ずしも生息地に近い場所での気温でないことは注意しておく必要がある.
気象台は次の通り.福岡,熊本,鹿児島,松山,徳島,高松,下関,広島,岡山,神戸,大阪,京都,和歌山,松江,鳥取,津,名古屋,静岡,横浜,東京,千葉.第二次分布拡大期で,一つの目安となる6℃(図22参照)を恒常的に下回っている地域は,京都,奈良,鳥取,松江,名古屋で,山陰,盆地など冬季に気温が下がりやすい地域である.飛び抜けて高いのは鹿児島.
これだけを見ると,南方種の分布拡大が冬季気温上昇で説明できるのではないかという印象が強くなりそうです.そこで,温暖化とは異なる視点で分布拡大(北上)を解釈する議論を一つ検討しておきましょう.
※13.最近日本において各地の平均気温が上昇していることは観測データから明らかで,これは事実である.しかし地球温暖化が進行しているかどうかに関しては,さまざまな議論があり,私には答えられない.したがって,ここでは「温暖化」という語を,最近進行している日本の気温上昇という意味で用いている.
渡辺(1989)は,琉球列島において分布拡大(北上)が著しい5種のトンボ(アオビタイトンボ,ヒメキトンボ,ベニトンボ※14,コシアキトンボ,オオキイロトンボ)を選び,それらの分布拡大について議論しています.これらのトンボの共通点としては,ベニトンボなどは流水にも生活していますから一概にいえませんが,いずれも止水性の種(池にすむ種)であるということがあげられます.かつての琉球列島の小さな島々には,目立った池沼は決して多くはなかったはずで,水環境としては流水環境,それも森林に囲まれたような環境が主体であったと思われます.それが近年のダム建設に代表される人為的な明るい止水域の創出によって,これらのトンボの生活領域が誕生し(新たなニッチの出現),定着をなしえたと渡辺氏は解釈しています.
わたしは渡辺氏との私信の中で,この北上を温暖化の枠組みでとらえられないかを検討しましたが,ほとんど否定的でした.図19は石垣市,那覇市,名瀬市,鹿児島県枕崎の1972~1992年までの冬季平均気温の変化に,上記の種が発見された時期を書き入れたものです.1977年沖縄本島で記録される(渡辺,1977)まで,日本国内では大東島でしか見ることのできなかったアオビタイトンボは,わずか9年後の1986年に鹿児島県加世田市(杉村・宮畑,1986)で見つかっていますが,枕崎と那覇の気温を比べて明らかなように,それを説明するだけの冬季平均気温の上昇はありません.
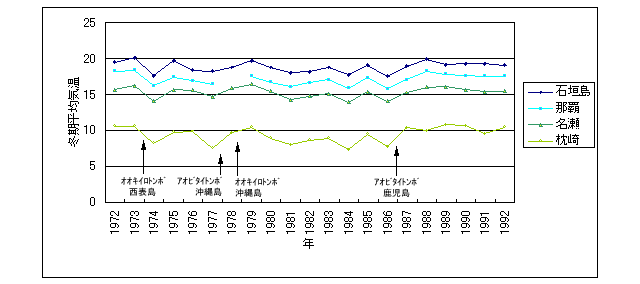
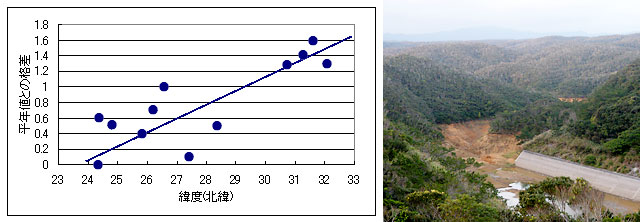
わたしは渡辺氏との私信の中で,この北上を温暖化の枠組みでとらえられないかを検討しましたが,ほとんど否定的でした.図19は石垣市,那覇市,名瀬市,鹿児島県枕崎の1972~1992年までの冬季平均気温の変化に,上記の種が発見された時期を書き入れたものです.1977年沖縄本島で記録される(渡辺,1977)まで,日本国内では大東島でしか見ることのできなかったアオビタイトンボは,わずか9年後の1986年に鹿児島県加世田市(杉村・宮畑,1986)で見つかっていますが,枕崎と那覇の気温を比べて明らかなように,それを説明するだけの冬季平均気温の上昇はありません.
図19.石垣市,那覇市,名瀬市,鹿児島県枕崎の1972~1992年までの冬季平均気温の変化.
図20は国内の各地の気象データから書いたもので,一般に冬季の平均気温の上昇は,緯度が高くなるほど大きくなる傾向があります.したがって,これら南方における各種の分布拡大は,温度上昇というよりは渡辺氏が指摘するように,開発行為の影響を受けたものと考える方が妥当だと考えられます.写真8は沖縄本島で造成されたダムで,こういった今までにない環境の出現が,新しい種を呼び込んでいるのでしょう.左:図20.「平年」と「1991-1992年にかけての冬(暖冬だった)」の平均気温の格差(12月から2月にかけて)と緯度の関係.数値は南西諸島から九州にかけての各気象台測候によるもの.地点は,西表島(N24.23.0),石垣島(N24.09.8),宮古島(N24.47.4),南大東島(N25.49.7),那覇(N26.12.2),名護(N26.35.4),沖永良部(N27.25.7),(名瀬(N28.22.6),種子島(N30.44.1),枕崎(N31.16.1),鹿児島(N31.34.4),阿久根(N32.01.5).直線は回帰させたものではなく,傾向を示しているだけである.見て分かるように,緯度が高いほど暖冬の程度は高くなる.
右:写真8.沖縄本島に造成されたダム.今までになかった広大な開水面を持つ止水域が誕生した.
しかしながら,私は,この議論はタイワンウチワヤンマには当てはまらない,と考えています.理由は次の通りです.タイワンウチワヤンマは止水性の種で,池沼で生活しています.タイワンウチワヤンマは昔から那覇市首里にある竜潭池で多く生息していることが知られていて,一般に開けた広い池沼で生活しているとされています.わたしは沖縄では,水田や,わずか3mX2mほどの防火用水桶でも幼虫を採集していますので,この種の繁殖可能な水域はかなり幅があるものと理解しています.右:写真8.沖縄本島に造成されたダム.今までになかった広大な開水面を持つ止水域が誕生した.
※14.ベニトンボはこの北上傾向が語られるよりずっと以前から鹿児島県南部に生息する個体群があることが知られていた.この個体群は沖縄の個体群とDNAの解析ではほとんど差はないという(尾園ら,2012).そして最近はタイワンウチワヤンマの後を追うように北上傾向が著しく,温暖化によって北上が推進されている可能性が高い.

写真9.タイワンウチワヤンマの見られる池の一つ.水生植物の有無や樹林の有無,また水際に対する要求など,特別な環境を必要とするわけではなく,ふつうのため池に見いだされる.
1980年代以降タイワンウチワヤンマが出現しはじめた瀬戸内海沿岸地域には,降水量の少なさから,非常に多くの灌漑用ため池があります.たとえば兵庫県は公称44,293カ所で全国一位(兵庫県農林水産部1998年調べ),以下全国上位県は,広島県(20,988),香川県(16,158),山口県(12,482),岡山県(10,284)と続きます(これらは1989年農林水産省調べ,以上兵庫水辺ネットワーク「ため池の自然」より引用).これらは歴史上相当古くから作られているものも多く,これらの水域の出現は決して今に起きたことではありません.したがって大量の止水域の出現と分布拡大の時期は一致しません.
それらに対して,温度の上昇は,空間的・時間的に,分布拡大とよく一致します.まず空間的に一致する例として,私の1992年の考察を紹介します.これは冬季平均気温※15の上昇と分布拡大を比較したもので,等温線の北上と,分布の北上を重ねたものです(図21).これを見ると,1987年以降,大阪湾地域では暖冬が続いて等温線が北へずれ,それに呼応するようにタイワンウチワヤンマの分布域が北へシフトしています.
タイワンウチワヤンマが温度に依存して北上しているなら,これらの種の現在の分布最前線とよく似た分布域を示しながら北上すると考えられます.図23は2000年頃の状況を示していますが,2020年現在山陰の方に分布拡大している状況はアオモンイトトンボのそれによく似ています.
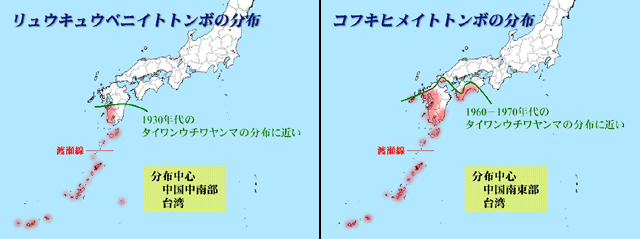
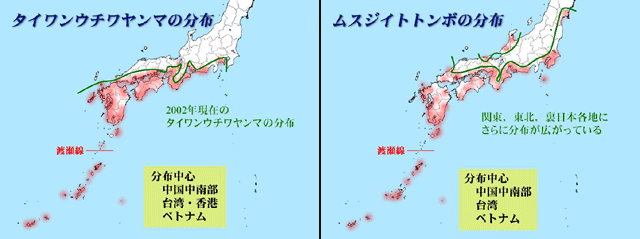
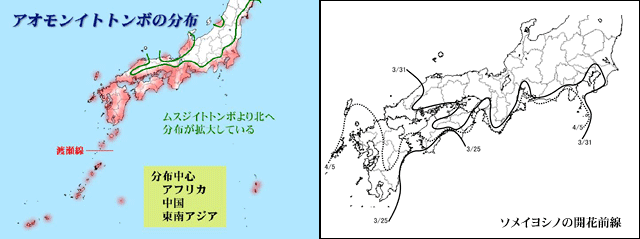
図21.等温線の北上と分布域の拡大.左が1987年より前のアメダス準平年値による冬季平均気温の等温線とタイワンウチワヤンマの分布,右が1987年以後の冬季平均気温の等温線と同種の分布である(青木,1992).
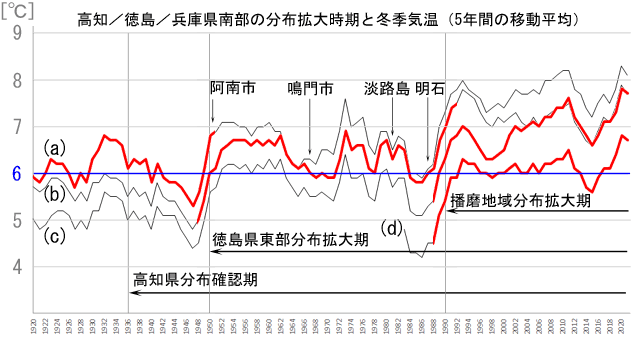
時間的に一致する例としては,青木(2021)で発表したグラフ(図22)をご覧ください.兵庫県の播磨地域へは,先に検討したように高知県から徳島県東部,淡路島を経て進入してきたと推定しています.各地域とも,この100年ほどの間に冬季気温が上昇しているのが分かりますが,高知県,徳島県東部,兵庫県本土側南部で分布を拡大した時期に,それぞれの冬季平均気温がほぼ同じ範囲になっているのが分かります.つまりおおよそ6℃(または少し下)を超える気温が安定的に続くようになったら,その地域内に分布が広がっています.図22.高知県,徳島県東部,兵庫県播磨地域で分布を拡大した時期とその時期の冬季平均気温.
(a) 高知地方気象台,(b) 徳島地方気象台,(c) 神戸海洋気象台/神戸地方気象台,(d) アメダス明石,それぞれの記録から見た冬季平均気温の変化と,それぞれの地域に分布拡大した時期を示す.いずれの地域においても,冬季平均気温が安定しておおよそ6℃(または少し下)を超えるぐらいになった時期に分布が拡大している.1950年代に兵庫県南部の冬季気温が6℃を超える時期が続いているが,この時期はまだ分布拡大が追いついていない(青木,2021を改変:青木(2021)図3キャプションで(d)をアメダス明石としているのは間違いで,(d)アメダス明石と神戸地方気象台の中間値が正しい).
タイワンウチワヤンマの現在の分布拡大が,温暖化によって引き起こされているとするならば,分布の最前線は温度によって決まっていると考えられます.そこで,温度に対する抵抗性の異なると思われる他種が日本本土でどのような分布をしているかを次に調べてみました.タイワンウチワヤンマと同様,東洋区つまり渡瀬線より南に分布中心があり,渡瀬線を越えて日本本土に入り込んでいるイトトンボ科の4種(リュウキュウベニイトトンボ,コフキヒメイトトンボ,ムスジイトトンボ,アオモンイトトンボ)を取り上げてみました(図23).これらは南方の種ですから分布最前線は温度によって決まっていると考えられ,かつ過去にすでに日本本土への分布拡大を「完了している種」といえます.分布域の違いは,おそらく温度に対する抵抗性の違いを反映しているのでしょう.(a) 高知地方気象台,(b) 徳島地方気象台,(c) 神戸海洋気象台/神戸地方気象台,(d) アメダス明石,それぞれの記録から見た冬季平均気温の変化と,それぞれの地域に分布拡大した時期を示す.いずれの地域においても,冬季平均気温が安定しておおよそ6℃(または少し下)を超えるぐらいになった時期に分布が拡大している.1950年代に兵庫県南部の冬季気温が6℃を超える時期が続いているが,この時期はまだ分布拡大が追いついていない(青木,2021を改変:青木(2021)図3キャプションで(d)をアメダス明石としているのは間違いで,(d)アメダス明石と神戸地方気象台の中間値が正しい).
タイワンウチワヤンマが温度に依存して北上しているなら,これらの種の現在の分布最前線とよく似た分布域を示しながら北上すると考えられます.図23は2000年頃の状況を示していますが,2020年現在山陰の方に分布拡大している状況はアオモンイトトンボのそれによく似ています.
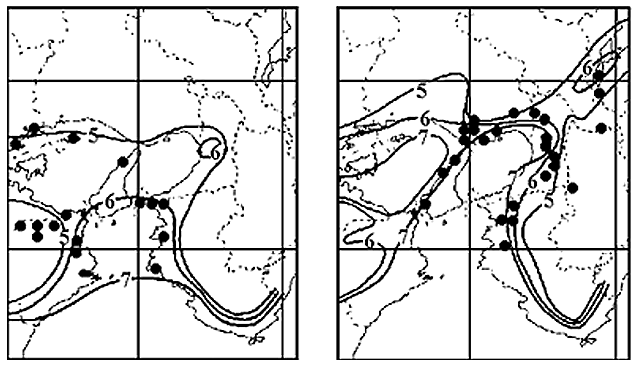
図23.温度に対する抵抗性が異なると考えられるイトトンボ科4種,リュウキュウベニイトトンボ,コフキヒメイトトンボ,ムスジイトトンボ,アオモンイトトンボの分布最前線(緑色)と,タイワンウチワヤンマの2000年頃の分布最前線(緑色).
タイワンウチワヤンマの過去の分布最前線(図9参照)と,リュウキュウベニイトトンボ,コフキヒメイトトンボの分布最前線がよく一致している.また今後温暖化が進むと,おそらく,ムスジイトトンボやアオモンイトトンボのような分布になっていくかもしれない.タイワンウチワヤンマはこれらのちょうど中間の位置に分布最前線があることが分かる.なお,温度に依存する前線として有名なソメイヨシノの開花前線も資料として掲げた.これらのトンボの分布前線と重なることが見て取れる.
このように,冬季気温と分布拡大の時間的・空間的相関性は,かなりよいといえるでしょう.以上から,私は,「今」タイワンウチワヤンマが北へ分布を拡大しているいちばん大きな原因は,最近の冬季気温上昇であると考えています.そしてこの最近の日本の気温上昇が地球温暖化によるものだとしたら,本種の北上は,地球温暖化が小さな昆虫に与えている影響を示す一つの例になるといえるでしょう.
タイワンウチワヤンマの過去の分布最前線(図9参照)と,リュウキュウベニイトトンボ,コフキヒメイトトンボの分布最前線がよく一致している.また今後温暖化が進むと,おそらく,ムスジイトトンボやアオモンイトトンボのような分布になっていくかもしれない.タイワンウチワヤンマはこれらのちょうど中間の位置に分布最前線があることが分かる.なお,温度に依存する前線として有名なソメイヨシノの開花前線も資料として掲げた.これらのトンボの分布前線と重なることが見て取れる.
※15.平均気温を使ったのは,越冬幼虫が水温の影響を受けるのは瞬間的な最低気温よりは持続的な低温であると思われる.それは,冬季には発育の進行は停止していても摂食活動等の生存活動は行われているからである.したがって,平均気温の方が低温の影響を表すのによい指標であると考えられる.
温暖化? 北上するタイワンウチワヤンマ 改訂版
みなさんもタイワンウチワヤンマを探してください

写真10.左:タイワンウチワヤンマの成虫.右:ウチワヤンマの成虫
腹部の先端にあるうちわ上の付属物内に黄色の紋があればウチワヤンマである.タイワンウチワヤンマにはそれがない.
腹部の先端にあるうちわ上の付属物内に黄色の紋があればウチワヤンマである.タイワンウチワヤンマにはそれがない.
さて,タイワンウチワヤンマは上の左の写真のようなトンボです.7月から9月下旬にかけて,ため池の,水から突き出た棒の先端に水平に止まっている姿がよく見られます.大きさは,腹長が50mm程度の中型のトンボです.「ヤンマ」という名前がついていますが「ヤンマ」の仲間ではなく,「サナエトンボ」の仲間です.よく似たトンボに,ウチワヤンマというのがいます(写真10右).これはかなり北の方までいるトンボで,北上とは関係ありません.腹部の先端のウチワ状のひらひらが特徴で,これはタイワンウチワヤンマにもありますが,ウチワヤンマでは内部に黄色い部分があるのが特徴です.写真で見比べてみてください.
また7月には池で抜け殻が見つかることがあります.これがあると飛来したのではなく,そこで冬を越したことになりますから,証拠としてはより重要です.写真11にウチワヤンマとタイワンウチワヤンマの羽化殻の写真を掲げておきました.ぜひ皆さんもタイワンウチワヤンマを探してみてください.特に,兵庫県,京都府,福井県の日本海側,千葉県,茨城県太平洋側などがホットスポットになると思います.
また7月には池で抜け殻が見つかることがあります.これがあると飛来したのではなく,そこで冬を越したことになりますから,証拠としてはより重要です.写真11にウチワヤンマとタイワンウチワヤンマの羽化殻の写真を掲げておきました.ぜひ皆さんもタイワンウチワヤンマを探してみてください.特に,兵庫県,京都府,福井県の日本海側,千葉県,茨城県太平洋側などがホットスポットになると思います.

写真11.羽化殻.左:タイワンウチワヤンマ,右:ウチワヤンマ.
参考文献
青木典司,1991.神戸市のサナエトンボ調査.Gracile 46:13-18.
青木典司,1992.タイワンウチワヤンマ幼虫の神戸市での越冬記録.Tombo 35(1/4):47-50.
Aoki, T., 1997. Northward expansion of Ictinogomphus pertinax (Selys) in eastern Shikoku and western Kinki Districts, Japan (Anisoptera: Gomphidae). Odonatologica, 26(2):121-133.
青木典司,2000.記録から見た国内におけるタイワンウチワヤンマの分布拡大の様相.Tombo 42(1/4):15-22.
青木典司,2021.北上するタイワンウチワヤンマ.昆虫と自然 56(12):12-17.
青木典司・二宗誠治,1998.兵庫県西部にタイワンウチワヤンマをさがして.Sympetrum Hyogo 5:16-19.
青野孝昭,2011.岡山県内タイワンウチワヤンマの分布調査報告.すずむし(146):43-46.
青山幹男・山本晃弘・福永みちる・中坪孝之,2014.広島大学東広島キャンパスのトンボ相.広島大学総合博物館研究報告6:51-60.
朝比奈正二郎,1957.日本の蜻蛉資料[11].新昆虫 10(6):51-58.
東 輝弥,2010.兵庫県のトンボ分布目録.Sympetrum Hyogo (12).
東 輝弥・青木典司,1999.西へ動き出した?,兵庫県南部のタイワンウチワヤンマ.Sympetrum Hyogo 6:2.
安達隆昌・藤原弘史・秋山美文,1996.広島県産トンボの分布記録.比和科学博物館研究報告 34:111-122+2 pls.
乾風 登,1981.湯浅町でタイワンウチワヤンマの生息を確認.Kinokuni 20:34.
乾風 登,2011.県内で初めてのベニトンボ.ことのうら (100):12.
乾風 登,1991.田辺市で見つかったタイワンウチワヤンマとその生息地.くろしお 10:25-26. 南紀生物同好会会報.
乾風登・高松勉,1986.和歌山県の日高町と美浜町でのタイワンウチワヤンマ.Kinokuni 30:12.
Corbet, P. S., 1962. A Biology of Dragonflies. H. F. & G. Witherby ltd., GB.
Corbet, P. S., 1999. Dragonflies: behavior and ecology of Odonata. Comstok, Cornell Uviv. Press, New York.
福井順治・加藤哲男,1997.1996年のタイワンウチワヤンマの分布調査.駿河の昆虫 178:5001-5006.
藤本勝行,1991.滋賀県におけるトンボの小知見.Aeschna 25:22-23.
藤原弘史・安達隆昌,1989.豊田郡大崎上島でのトンボ採集記録.広島虫の会会報 28:46.
Gurney, S. C. William, Philip H. Crowley, & Roger M. Nisbet, 1992. Locking life-cycles onto seasons: Circle-map models of population dynamics and local adaptation. J. Math. Biol. 30:251-279.
浜田 康・井上 清,1985.日本産トンボ大図鑑.講談社.
久川 健,1994.北九州市とその付近のトンボ(上),-1993年の採集記録-.わたしたちの自然史 49:19-23.
久川 健,1991.奈良県五條市付近で注目すべきトンボの採集記録.Gracile 45:32-35.
平井雅男,1968.小松島市のトンボ類.総合学術調査報告 小松島 郷土研究発表会紀要 14:49-55. 阿波学会.
平井雅男,1971.吉野川中流地方のトンボ類.総合学術調査報告 麻植パイロット開拓地帯 郷土研究発表会紀要 17:28-34.阿波学会.
平井雅男,1973.脇町およびその周辺のトンボ類.総合学術調査報告 脇町及びその周辺 郷土研究発表会紀要 19:29-33.阿波学会.
平井雅男,1980.池田町のトンボ類.総合学術調査報告 池田町 郷土研究発表会紀要 26:23-30. 阿波学会.
平松恒彦,1979.中津・宇佐平野のため池の水生動物.大分県自然環境保全地域候補地調査報告書(県北地区):21-24.大分県.
池崎善博,1961.長崎市付近のトンボ.染色体 15:23-25. 長崎高校生物部.
池崎善博,1977.長崎県松浦市でタイワンウチワヤンマの脱皮殻を採集.Tombo 20(1/4):22.
池田 寛,1967.山口県におけるタイワンウチワヤンマの記録.山口県の自然 (18):32. 山口県立博物館.
池田 寛,1969.山口県のタイワンウチワヤンマ 幼虫の発見と県下の新産地について.山口県の自然 (22):5-6. 山口県立博物館.
池田 寛,1988.トンボ目 ODONATA.山口県の昆虫:7-15.山口県立博物館編,197+32pls.
石川 一,1982.香川県産のタイワンウチワヤンマとオオキトンボ.昆虫と自然 17(2):33.
石川 一,1983.タイワンウチワヤンマ香川県西部に定着か(?).昆虫と自然 18(14):12.
石川 一,2002.神奈川県でタイワンウチワヤンマ採集.Tombo 45(1/4):40.
石川 一・伊藤信男,2016.東京に出現したタイワンウチワヤンマ.神奈川虫報(189):97-98.
石田昇三・石田勝義,小島圭三・杉村光俊,1988.日本産トンボ幼虫・成虫検索図説.東海大学出版会.
市場利哉,1973b.佐賀県トンボ類分布資料(1).佐賀の昆虫 (1):33-34.
井上 清,1988.愛媛県大島のトンボ採集記録.Tombo 31(1/4):42-45.
井上 清・相浦正信,1995.壱岐のトンボ分布記録(第1報).Tombo 38(1/4):66.
井上 清・小川雅由,1991.タイワンウチワヤンマを西宮市内で発見.Gracile 45:20-22.
片谷直治,2017.Ictinogomphus rapax (Rambur, 1842)とI. pertinax (Selys, 1854)(タイワンウチワヤンマ)の形態比較と2種の分布.Aeschna (53):1-10.
苅部治紀,2013.酒匂平野のタイワンウチワヤンマの記録.神奈川虫報(180):40-41.
苅部治紀・小浜継雄,1990.北大東島のトンボ相.Tombo 33(1/4):24-26.
岸 一弘,2012.神奈川県小田原市におけるタイワンウチワヤンマの記録.月刊むし (499):45.
楠博幸・桑田一男,1979.愛媛県のトンボ.愛媛県立博物館研究報告 10.35pp+2pls.
窪田聖一,1982.タイワンウチワヤンマ愛媛県松山市にて採集.月刊むし 140:33.
倉敷市立自然史博物館,1994.倉敷のいきもの.第10回特別展解説資料.
神戸海洋気象台,1993-1994.気象月報.
舛田誠二,1959.タイワンウチワヤンマ熊本市付近に多産す.熊本昆虫同好会報 5(2):24.
松木和雄,1993.倉品治男氏採集の福岡県苅田市のトンボ.九州虫の会会誌 13:33-34.
松木和雄・藤田慎一・松比良邦彦・宮川続・吉崎一章・久川健・倉品治男・今村久雄・今村浩継・渡辺庸子,1994.鹿児島県川内市周辺のトンボ類.九州虫の会会誌 15:1-11.
松田 勲,1987.兵庫県家島諸島男鹿島でタイワンウチワヤンマを採集.Gracile 37:7-8.
松本導男,2001.高知県南国市のタイワンウチワヤンマ羽化記録.Gracile (63):11-15.
宮本詔子・宮本聡史,2008.益田市でタイワンウチワヤンマの成虫と羽化殻を採集.すかしば (56):49-51.
村木明雄,1980.和歌山県加太市でタイワンウチワヤンマを発見.Gracile 27:6-7.
永井あつし,1974.宮崎県産トンボ資料.タテハモドキ 9:8-9.
中岡芳彦, 1996. 静岡県においてタイワンウチワヤンマの羽化殻を多数採集.Tombo 39(1/4):47.
長瀬 翔・永幡嘉之,2022.鳥取県東部でタイワンウチワヤンマを採集.すかしば (59):38.
長嶺邦雄,1964.竜潭池における不均翅亜目(蜻蛉目)の羽化の観察.このは会会報 6(1):1-18.
中村重久,1957.昆虫覚え書(Ⅱ).げんせい 5(2):45-48.
Naraoka, H., 1987. Studies on the ecology of Cercion sieboldii (Selys) in the Aomori Prefecture, northern Japan. I. Life history and larval regulation (Zygoptera: Coenagrionidae). Odonatologica 16: 261-272.
成田茂生,1988.三重県におけるタイワンウチワヤンマの生態.Aeschna (21):16-20.
二宗誠治,1988.タイワンウチワヤンマ,明石公園で発見.Gracile 39:34.
Norling, U., 1984. Life history patterns in the northern expansion of dragonflies. Adv. Odonatol. 2:127-156.
尾園暁・桜谷保之,2005.奈良県のトンボ相-1998年~2003年の記録-.近畿大学農学部紀要 (38):71-155.
尾花 茂・津田 滋・井上 清・冨士原芳久・新村捷助,1979.「南西諸島飛び歩き」の採集品,幼虫飼育および現地観察の記録.Gracile (25):5-9.
大浜祥治,2009.2008年,島根県のタイワンウチワヤンマ分布状況調査.すかしば (57):27-28.
大浜祥治,2012.2009-2011年,山陰地方沿岸部におけるタイワンウチワヤンマ分布状況調査.すかしば(59):30-31.
大浜祥治,2013.鳥取県初記録のタイワンウチワヤンマを境港市で採集.すかしば(60):42.
佐賀北高校生物部昆虫班,1967.上無津呂・羽金山一帯の昆虫.生物記 5:15-28. 佐賀県立佐賀北高等学校生物部.
佐々木重治,1936.タイワンウチハトンボ Ictinus fallax Selys 土佐に採集せらる.昆虫界 4(26):235-237.
笹本彰彦,1998.奈良盆地でのタイワンウチワヤンマの記録.Gracile 59:28-29.
佐藤利明,1980.大分県蜻蛉目録.自刊.
佐藤國康,1993.岡山県産タイワンウチワヤンマの古い記録.しぜんくらしき 5:10-11.
椎木昌則,1993.タイワンウチワヤンマを採集.広島虫の会会報 32:26.
下野伸司・山口和洋,1991.紀南地方のトンボ.ひらくら 35(6):81-87.
Siva-Jothy, M.T., 1997. Odonate ejaculate structure and mating systems. Odonatologica 26(4):415-437.
新村捷介,1989.淀川ワンドでタイワンウチワヤンマを採集.Gracile 42:43.
杉村光俊,1981.トンボ数種の産卵に関する知見と考察(その2).Tombo 23(1/4):35-38.
杉村光俊・宮畑年弘,1986.アオビタイトンボの九州本土における記録.Tombo 29(3/4):112-113.
杉村光俊・山崎憲男・鵜飼貞行,1989.トンボ王国ガイド.財団法人トンボと自然を考える会.
杉村光俊・石田昇三・小島圭三・石田勝義・青木典司,1999.原色日本トンボ幼虫・成虫大図鑑.北海道大学図書刊行会.
Selys-Longchamps, 1888. Odonates recueillis aux iles Loo-Choo par feu Pryer. C. r. Sic. Ent. Belg., 32: 1-6.
互井賢二・芝原達也,2019.タイワンウチワヤンマ千葉県で発見される.房総の昆虫(65):38-39.
高木真人,1986a.香川県東讃地域のトンボ相.香川県自然環境保全指標策定調査報告書(香川県東讃地域):193-209.
高木真人,1987b.小豆島のトンボ相.香川県自然環境保全指標策定調査報告書(香川県小豆島地域):139-146.
高松 勉,1993.紀伊半島南東部和歌山県でのタイワンウチワヤンマの初記録.Gracile 50:17.
高松 勉,1988.タイワンウチワヤンマの新産地.Kinokuni 33:22.
竹内 勉,1976.近畿地方より得られた珍しいトンボ3種について.Gracile 20:19-20.
田中 稔,1982.タイワンウチワヤンマとウチワヤンマ混棲す.Parnassius 26:12.
田端 修,2001.京都府南部でタイワンウチワヤンマを記録.Gracile(63):23-24.
津田滋・山本哲央,1988.大阪府でタイワンウチワヤンマを採集.Gracile 39:13-14.
冨田靖男・市橋甫,1971.タイワンウチワヤンマ鈴島に産す.ひらくら 15(11):81-82.
蜻蛉研究会編,1998.滋賀県のトンボ.琵琶湖博物館研究調査報告 10.283pp.滋賀県
鵜殿清文,2000.愛知県でタイワンウチワヤンマを採集.月刊むし 347:44-45.
渡辺賢一,1977.石垣島産蜻蛉4種について.Tombo 20(1/4):27-28.
渡辺賢一・小浜継雄,1985.沖縄のトンボ.70pp+7pls. 沖縄県立博物館,那覇.
渡辺賢一,1989.琉球列島における分布拡大の蜻蛉種について.Tombo 32(1/4):54-56.
山崎 晃,1992.タイワンウチワヤンマ玉島に定着.しぜんくらしき 1:11.
山口県立山口博物館編,2006.山口県のトンボ.
吉田雅澄・八木孝彦,2012.東海地方3県におけるタイワンウチワヤンマの分布拡大.Aeschna(48):37-41.
吉田雅澄・八木孝彦,2013.続・東海地方3県におけるタイワンウチワヤンマの分布拡大.Aeschna(49):29-32.
吉田雅澄・八木孝彦・鵜殿清文,2014.東海地方3県におけるタイワンウチワヤンマの分布拡大,第3報.Aeschna (50):73-77.
吉田雅澄・八木孝彦・鵜殿清文,2015.東海地方3県におけるタイワンウチワヤンマの分布拡大,第4報.Aeschna (51):21-24.
吉田雅澄・鵜殿清文・八木孝彦,2016.東海地方3県におけるタイワンウチワヤンマの分布拡大,第5報.Aeschna (52):33-37.
吉田雅澄・鵜殿清文・八木孝彦,2017.東海地方3県におけるタイワンウチワヤンマの分布拡大,第6報.Aeschna (53):59-63.